Article publié en 1970 dans la Revue Forestière Française, n°2, p 131 - (version .pdf - 287k)
La notion de niveaux d'énergie et son intérêt en sylviculture
par Louis ROUSSEL
(Les chiffres placés entre parenthèses dans le texte renvoient à la bibliographie.)
I - GENERALITES SUR LE ROLE
DE L'ENERGIE NATURELLE DANS LA PRODUCTION LIGNEUSE
On a beaucoup écrit, ces dernières années,
sur " l'usine forêt " et sur l' " arbre machine "
qui, grâce au gaz carbonique de l'air, à l'eau et aux matières
minérales du sol, fabriquent, d'une façon plus ou moins continue,
des substances ligneuses diverses (en très grosse majorité du
type " polyholosides "), en utilisant l'énergie naturelle fournie
par le soleil. D'une production théorique de 3000 tonnes de matières
sèches par hectare et par an (si l'on admet que la totalité des
1013 petites calories reçues, chaque année, par un
hectare de surfaces boisées, dans la partie Nord-Est de notre pays, pourraient
être utilisées à la production ligneuse), on tombe, pratiquement,
à 5 ou 6 tonnes seulement de matières sèches, extraites
annuellement, souvent moins, rarement davantage. Le rendement énergétique
brut de " l'usine forêt " ressort donc à 0,2 % environ
(1). En agriculture, le même type de raisonnement conduit à un
chiffre analogue, soit 0,2 % (2).
Mais, on peut se borner à considérer l'énergie
incidente pendant la période de végétation la plus active
(en gros, d'avril à septembre ou octobre), ou même la seule énergie
réellement absorbée, pendant cette période, par les appareils
foliacés (déduction faite de ce qui est renvoyé vers le
ciel, ou albédo, de ce qui est retenu par les branchages et par les troncs,
et de ce qui est transmis au sol). Dans ce dernier cas, et en évaluant
non la masse ligneuse extraite par les récoltes annuelles, mais la totalité
de la " biomasse " produite (bois du tronc et des branches, feuillages,
racines), A. Galoux obtient des chiffres très supérieurs de rendement
énergétique, et qui se rapprochent de 6 % (3). En 1968, du reste,
cet auteur a avancé des données encore plus favorables,
Le lecteur aura facilement compris qu'il n'y a aucune discordance
profonde entre ces divers éléments. Seule change la façon
de présenter le problème.
De toutes façons, on ne doute plus guère actuellement, sauf chez quelques partisans attardés des théories de l'humus, que cette façon d'envisager la production forestière sous son aspect énergétique, ne soit tout à fait justifiée, et que les considérations théoriques qu'elle suscite ne puissent ouvrir des perspectives nouvelles, et des plus intéressantes pour les sylviculteurs. Le problème est, cependant, des plus vastes et des plus complexes, et l'on se bornera, dans les lignes qui vont suivre, à ne mettre en relief que certaines des idées de base les mieux établies, et, aussi, à signaler les lacunes ou les incertitudes qui freinent encore les progrès des chercheurs.
Il - REPARTITION VERTICALE DU RAYONNEMENT NATUREL
Cas des peuplements homogènes
En photométrie, le flux énergétique
ou lumineux reçu à un niveau quelconque (I) d'un milieu bien homogène,
s'exprime par la relation classique :
F (I) = F (o). e-aI
Dans cette formule, F (I) représente, comme
il est dit, le flux reçu au niveau (I). F (o) est le flux incident,
à son entrée dans le milieu homogène, e est la base
de logarithmes népériens, et a le coefficient d'extinction,
englobant l'ensemble des coefficients de diffusion, de réflexion et d'absorption.
Ch. Perrin de Brichambaut, dans un récent et important ouvrage (4), indique
la façon dont cette relation est établie. L'exposant aI
en particulier, est relié à la densité optique d,
notion classique également, par l'égalité d = 0,434
aI.
Dans une formation végétale bien homogène,
on adopte, en général un type de relation voisin, aussi bien dans
les cultures (5) que dans les forêts (6).
E (f) = E (o). e- Kf
E (f) = énergie (ou plus exactement "
flux énergétique ") reçue par unité de
surface au niveau (f).
E (o) = énergie incidente (flux énergétique)
reçue par unité de surface horizontale, au sommet de la végétation
(déduction faite de l'albédo, c'est-à-dire de la proportion
d'énergie renvoyée vers le ciel, par les cimes).
e = base des logarithmes népériens.
K = coefficient d'extinction des radiations.
f = indice foliaire partiel : surface des feuilles comprises entre
le niveau f et le sommet de la formation végétale, par
unité de surface du sol.
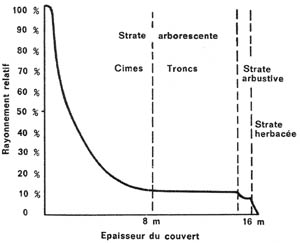
On remarquera que cette dernière formule diffère de la première sur certains points (introduction de l'albédo et de l'indice foliaire, remplaçant l'épaisseur du milieu homogène), mais, que d'une façon générale, il existe une grande similitude entre les deux. Par ailleurs, il est très probable que, dans la seconde formule, le coefficient d'extinction K varie avec le type de temps (plus élevé par temps ensoleillé, plus faible par temps couvert). La figure 1 représente l'allure de la courbe théorique reliant le flux énergétique à l'épaisseur du couvert, telle qu'elle a été établie par J. Grulois dans un travail récent (6).
Expérimentalement, on constate que le milieu
forestier se comporte, sous un type de temps déterminé, d'une
façon assez conforme à la théorie. Les figures 2 et 3,
présentées d'une façon un peu différente de celle
adoptée pour la figure 1, résultent de mesures de rayonnement
effectuées, en continu, ces dernières années, pendant des
périodes variables, dans une jeune plantation résineuse dense,
et dans une chênaie-hêtraie à charme, d'un type " taillis-sous-futaie
vieilli ", et assez homogène. Le niveau d'énergie N (o) est
fixé, pour éviter tout ombragement latéral de certaines
cimes, d'une hauteur parfois irrégulière, à quelques mètres
au-dessus de la surface des peuplements, sans déduction cette fois de
l'albédo, Les niveaux N (1), N (2), N (3) représentent, respectivement
50 %, 25 %, 12,5 %... de l'énergie reçue au niveau N (o). Pour
fixer les idées, on indiquera qu'au niveau N (o), et dans la région
Nord-Est de l'Europe occidentale, l'éclairement énergétique
est, pour une année complète, voisin de :
274 langleys (ly), soit 274 petites calories par centimètre
carré de surface horizontale par jour moyen ou 2637 Kilowatts
(kW) par hectare (*), en ne comptant que les heures de jour.
Pendant les six mois de végétation la plus
active (d'avril à septembre), les chiffres ci-dessus deviennent :
400 langleys par jour moyen ou 3850 kW / hectare (*), en
ne comptant que les heures de jour.
La correspondance entre les éclairements, énergétiques et lumineux, est assez difficile à établir. L'énergie solaire, en effet, comprend des rayons de longueurs d'onde variées (de 0,3 à 2,5 ou 3 µ), alors que la lumière, partie visible du rayonnement, ne s'étend que de 0,4 à 0,7 µ environ. MAURAIN, en 1937, se basant sur les travaux classiques de KIMBALL retient la correspondance suivante : 1 langley par minute = 72000 lux. DOGNIAUX (1960) en Belgique, à la suite d'observations de longue durée, admet un chiffre très voisin : soit 1 langley par minutes = 70000 lux. En Autriche, TRANQUILLINI (1960) donne des éléments un peu supérieurs : 1 langley par minute = 75000 lux, mais, dans les hautes régions des Alpes centrales, TURNER (1958) arrive à 1 langley par minute = 96000 lux. Au contraire, très récemment, GRULOIS et VYNCKE (1969), dans les Ardennes belges, trouvent que 1 langley par minute équivaut seulement à 53 500 lux environ, en plein découvert, beaucoup moins dans les sous-bois feuillus, en raison de l'émission, dans ce milieu, d'une forte quantité de rayons infrarouges par la face inférieure des feuilles. Cependant, en s'en tenant aux données moyennes (MAURAIN, KIMBALL, DOGNIAUX, TRANQUILLINI), on peut calculer que l'éclairement moyen annuel, en plein découvert, est de l'ordre de 27 000 lux, et, pendant la période de végétation la plus active, de l'ordre de 34 000 lux, avec, évidemment, de fortes variations de part et d'autre de ces chiffres.
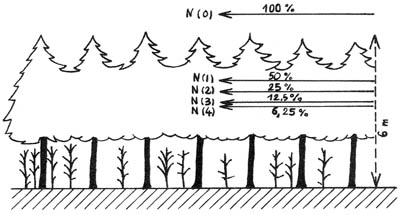
Figure 2. - Situation approximative des niveaux d'énergie ans
une jeune plantation d'épicéa (d'après BAUMGARTNER -1956)
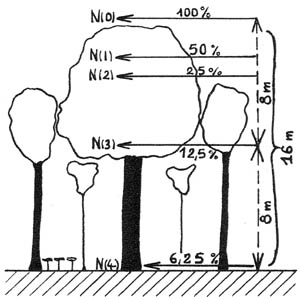
Figure 3. - Situation approximative des niveaux d'énergie dans une chênaie-hêtraie à charme (d'après GALOUX - 1968 et GRULOIS - 1967).
Cas des ouvertures de formes diverses
La relation reproduite plus haut ne s'applique évidemment pas quand le peuplement devient discontinu, donc très hétérogène. Dans ce cas, et à défaut de données expérimentales assez nombreuses qui font encore défaut, on peut utiliser la méthode graphique dite du " cercle d'illumination totale " (7).
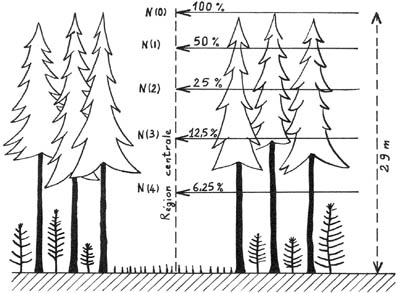
Figure 4. - Situation approximative
des niveaux d'énergie dans la région centrale d 'une petite trouée
(Méthode théorique ROUSSEL 1962).
La figure 4 indique comment l'énergie naturelle se répartirait verticalement, dans la région centrale d'une " petite " trouée circulaire, (dont le diamètre est égal à la moitié de la hauteur des arbres du peuplement voisin, supposé par ailleurs très dense), mais, cette fois, pendant toute l'année, à une latitude nord voisine de 45-50°. Sous cette dernière réserve, les niveaux d'énergie sont choisis de la façon indiquée au paragraphe précédent, et les valeurs absolues mentionnées sont définies de la même façon.
III - CONSIDERATIONS GENERALES SUR L'UTILISATION
DES NIVEAUX D'ENERGIE EN SYLVICULTURE
Les figures 1, 2, 3 et 4 montrent, avec évidence,
qu'il existe des niveaux d'énergie différents dans le milieu forestier.
Le sylviculteur peut, par son action, intervenir, afin, surtout, de déplacer
vers le bas chacun de ces niveaux (en diminuant la densité des peuplements
homogènes, ou en élargissant les ouvertures). D'une façon
très simple, par exemple, on peut relever un tel genre de modification
dans le cas des coupes de régénération en futaie régulière
: pendant la majeure partie de la durée de la révolution, la zone
de forte absorption d'énergie sera maintenue dans la partie supérieure
des peuplements principaux ; puis, elle sera abaissée progressivement
(au cours des coupes successives) jusqu'au voisinage du niveau du sol pour y
assurer le développement des jeunes éléments, naturels
ou artificiels, destinés aux récoltes futures.
À l'inverse, mais bien plus lentement, le sylviculteur
peut faire remonter la zone de forte absorption du rayonnement, en laissant
croître des peuplements de forte densité.
Si chaque variété de chaque espèce
ligneuse avait un " coefficient d'utilisation de l'énergie naturelle
" absolument constant, quel que soit son âge, le niveau d'énergie
dans lequel elle se développe, et l'intensité de l'éclairement
qui règne au voisinage de ses aiguilles ou de ses feuilles, les choses
seraient relativement simples. C'est du reste cette hypothèse qui a servi,
et qui sert encore, assez fréquemment, pour des études théoriques
concernant les rapports entre l'énergie naturelle et la production ligneuse.
On pourrait ainsi facilement montrer :
- que la nature même du traitement (par exemple en
futaie régulière ou en futaie jardinée), n'a, pratiquement,
aucune influence sur la production globale. Pour une quantité déterminée
d'énergie absorbée par les cimes (par exemple, dans une sapinière,
avec un éclairement énergétique incident estimé
sur une année complète à 2 637 kW / ha (*), diminué
d'un albédo de 10 % et d'une transmission au sol de 5 %, il résulterait
une absorption par les cimes de 2 242 kW / ha (*)), la production d'une espèce
donnée serait toujours la même, dans un type de sol déterminé,
suffisamment approvisionné en eau et en matières minérales
dissoutes.
- que la production ligneuse varie en fonction de la densité
des cimes (donc de celle des peuplements), d'une façon analogue à
celle qui est établie expérimentalement (tables de production
allemandes, par exemple), et ce d'une façon des plus logiques (1).
- que certaines exigences spéciales, relatives à
la proportion idéale entre les divers types de peuplements, et imposées
par les aménagistes (en vue d'assurer une production régulière
dans le temps), déterminent, pour des espèces données,
le mode de traitement lui-même. Par exemple, une forêt mélangée
de sapins et d'épicéas, dans le Jura ou dans les Vosges, doit
fournir des bois exploitables à 140 ans. Sa surface étant, par
hypothèse de 140 hectares, on devrait régénérer,
chaque année 1 hectare.
Pour que les renaissances naturelles puissent se développer
d'une façon assez rapide pour pouvoir s'insérer dans la suite
des peuplements, il est nécessaire que les jeunes sujets, pendant leurs
quinze ou vingt premières années, reçoivent approximativement,
de 900 à 1300 kW / ha (*). Cette exigence résulte de nombreuses
observations photobiologiques (1).
On peut envisager de recruter, chaque année, cet
hectare de régénérations, sous forme de " petites
trouées " (d'un diamètre égal à la moitié
de la hauteur des grands arbres sur pied). Avec des arbres de 30 mètres
de hauteur totale, ceci conduirait à 50 trouées de 2 ares, ne
donnant, au total, aux régénérations installées,
que 80 à 100 kW / ha (*).
Si l'on pratique des " trouées normales ",
l'apport de rayonnement atteindra 250 kW / ha (*) et sera encore insuffisant.
Avec de " grandes trouées ", d'un diamètre égal
à deux fois la hauteur des grands arbres sur pied, on se rapprochera
de l'optimum, avec 800 kW / ha (*). Mais ce n'est qu'avec de " très
grandes trouées " (d'un diamètre égal à quatre
fois la hauteur des grands arbres sur pied) que l'on sera assuré d'un
développement convenable des régénérations. Or,
avec des arbres de 30 mètres de hauteur, la " très grande
trouée ", représente un hectare. On est donc, dans le
cas envisagé, orienté vers la coupe à blanc, chaque année,
d'un hectare de peuplement ; ceci n'est plus exactement du jardinage, mais plutôt
de la futaie régulière.
Bien entendu, il n'est nullement impossible de retenir,
même avec l'épicéa pur, un traitement en futaie jardinée,
en pratiquant des " trouées normales ". Mais la croissance
des jeunes sujets sera ainsi ralentie, et leur incorporation dans la suite des
peuplements rendue plus difficile. Il faudra abandonner certaines des données
initiales : rapport soutenu, ou bien âge d'exploitation. Car ces diverses
considérations constituent un système " contraignant "
auquel Il est difficile de se soustraire - les aménagistes le savent
bien.
Dans le cadre de la conception initiale (chaque variété
de chaque espèce a un coefficient d'utilisation de l'énergie constant
quelque soit son âge, le niveau dans lequel elle se développe et
l'intensité de l'éclairement qu'elle reçoit), on peut imaginer
des types de futaies étagées (produisant surtout du bois de râperie
par exemple) à utilisation optimale des radiations naturelles : entre
les niveaux 0 et 1 seraient situées les cimes de pin ( à bois
pou coloré), entre les niveaux 1 et 2, celles d'épicéa,
et entre les niveaux 2 et 3 celles de sapin. Cette notion a, du reste, été
largement utilisée déjà par les sylviculteurs, mais sans
références de base bien précises.
On le répète : ce type de raisonnement paraît logique, il a été adopté par divers théoriciens à titre de première approximation, et l'auteur de ces lignes s'y est, parfois, lui-même conformé.
IV - COMPORTEMENT REEL DES ESPECES LIGNEUSES
Malheureusement pour les amateurs de notions simples, les
réactions des diverses espèces ligneuses paraissent un peu plus
nuancées. Il faut reconnaître, du reste, qu'elles sont encore assez
mal déterminées dans tous leurs multiples détails, Toutefois,
dans certains cas, des études très poussées ont déjà
été effectuées (grâce surtout aux appareils à
absorption de rayons infrarouges : URAS ou IRGA), notamment par les chercheurs
de l'Ecole d'Innsbruck (8 et 9). Le hêtre et l'épicéa, en
particulier, commencent à être assez bien connus en ce qui concerne
le rythme de leur photosynthèse en fonction de la lumière et de
la température, de l'âge des sujets, et de la position des appareils
foliacés dans les cimes. On peut calculer, dans divers cas, un "
coefficient d'utilisation de la lumière ", caractéristique
de l'activité photosynthétique des aiguilles et des feuilles,
dont la prise en considération semble intéressante. Le calcul
repose en effet sur des données expérimentales très précises
: la quantité de CO2, effectivement assimilée, dans
chaque cas, par unité de temps (une heure en général).
Si l'on prend comme base de référence 1 le " coefficient
d'utilisation " de l'appareil foliacé de pleine lumière,
sous 10000 lux, par unité de poids sec et dans la région de la
température optimale, on obtient ce qui suit :
|
Picea excelsa
|
Coefficient d'utilisation
|
|||
|
{
|
sous 10000 lux
|
1
|
||
|
Rameaux de lumière
|
sous 20000 lux
|
0,72
|
||
|
sous 30000 lux
|
0,54
|
|||
|
{
|
sous 10000 lux
|
1,43
|
||
|
Rameaux d'ombre
|
sous 20000 lux
|
0,88
|
||
|
sous 30000 lux
|
0,62
|
Ce qui signifie que le rameau de lumière absorbera deux fois plus de C02, pour une quantité de lumière déterminée, sous 10000 lux que sous 30000 lux (figure 5).
|
Fagus silvatica
|
Coefficient d'utilisation
|
|||
|
{
|
sous 10000 lux
|
1
|
||
|
Feuilles extérieures
(sommet des cimes) |
sous 20000 lux
|
0,60
|
||
|
sous 30000 lux
|
0,43
|
|||
|
{
|
sous 10000 lux
|
1,5
|
||
|
Feuilles extérieures
(base des cimes) |
sous 20 000 lux
|
1,1
|
||
|
sous 30000 lux
|
0,8
|
|||
|
Feuilles intérieures
|
{
|
sous 1000 lux
|
de 2 à
2,5
|
|
|
sous 10000 lux
|
0,5
|
Des relations analogues ont été trouvées
en 1965 par RETTER, mais les valeurs absolues sont un peu différentes,
en raison d'un perfectionnement apporté à la méthode de
travail (figure 6).
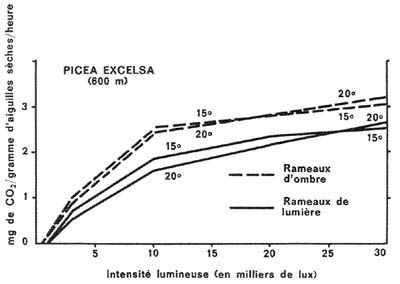
Figure 5. - Assimilation nette du gaz carbonique de l'air, en fonction
de la lumière, des aiguilles d'épicéa (intérieures
et extérieures des cimes), aux températures de 15° C et de
20° C (d'après PISEK et WINKLER-1959).
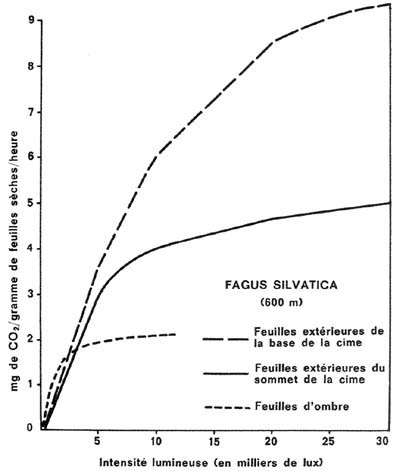
Figure 6. - Assimilation nette du
gaz carbonique de l'air, en fonction de la seule lumière, des feuilles
de hêtre extérieures des cimes (base et sommet) (d'après
PISEK & TRANQUILLINI - 1954) et des feuilles d'ombre typiques (d'après
BOYSEN-JENSEN 1932).
En somme, on peut relever, pour l'épicéa
et pour le hêtre :
- la supériorité, en ce qui concerne l'utilisation
de la lumière, des appareils foliacés développés
dans une lumière réduite, par rapport à ceux situés
en permanence dans la région la mieux éclairée des cimes.
Cette supériorité se maintient, pour l'épicéa,
aux éclairements élevés, mais s'atténue et disparaît
rapidement, dans le cas du hêtre.
- la " saturation " progressive, dans tous
les cas, des parenchymes chlorophylliens, lors de la majoration de l'éclairement
(par exemple, à partir de 10000 lux dans le cas du hêtre). Ceci
ne signifie nullement, du reste, que la feuille du hêtre ou l'aiguille
d'épicéa, développée dans une position déterminée
dans les cimes, n'assimile pas davantage, d'une façon absolue, sous 30
000 lux que sous 20 000 lux - mais que le rapport entre l'énergie
effectivement utilisée à assimiler le gaz carbonique de l'air,
et l'énergie incidente, diminue avec l'intensité de l'éclairement.
On remarquera, à ce propos, que les chiffres reproduits
ci-dessus ne font mention que de l'éclairement " lumineux
" (on l'a vu plus haut, il existe également un éclairement
" énergétique ") : ceci pour respecter intégralement
les données expérimentales publiées par les chercheurs.
Il y a cependant, on le sait, une correspondance entre ces deux éclairements,
mais les relations semblent complexes, Cependant, à titre de large approximation,
on pourrait retenir que l'éclairement de 10000 lux, pris comme base dans
le calcul du " coefficient d'utilisation de la lumière ", correspond
à un peu moins de 30 % de l'éclairement reçu en plein découvert.
Celui de 20000 lux correspond à 60 %, et celui de 30000 lux, à
90 % environ du même plein découvert. Ceci en considérant
la valeur moyenne enregistrée pendant les 5 à 6 mois de végétation
la plus active.
Le sylviculteur qui cherchera à étendre ces
notions rencontrera des difficultés sérieuses à trouver
des données aussi précises, pour toutes les espèces ligneuses
dont il envisage l'emploi. Certains éléments existent, d'une façon
éparse, dans de très nombreuses études publiées
récemment un peu partout dans le monde (les chercheurs japonais ont,
notamment, fait un gros effort dans cette direction). Aussi convient-il de signaler
un travail de synthèse important (quoique d'un volume réduit),
réalisé en 1969 par W. LARCHER (10), l'un des bons spécialistes
de ce genre de problème. Plus de 100 espèces d'arbres, situées
dans les régions les plus variées, sont ainsi sommairement analysées.
Voici quelques conclusions qui ressortent de l'examen des
multiples chiffres recueillis dans cette synthèse :
- de nombreux arbres résineux : Picea excelsa,
Picea glauca, Pinus cembra, Pinus resinosa, Abies balsamea, notamment, ont
non seulement un " coefficient d'utilisation de la lumière "
plus élevé pour leurs rameaux d'ombre que pour ceux de lumière,
mais la photosynthèse maximale nette des premiers, en lumière
de saturation spécifique, est voisine, et parfois supérieure,
à celle des rameaux de lumière, dans leur lumière de saturation
spécifique également. Les jeunes semis (et ceci est intéressant
au sujet de la conduite des opérations de régénération),
sont souvent plus actifs, du point de vue de leur nutrition carbonée,
quand ils se développent dans une lumière modérée,
que dans une lumière trop vive.
- par contre, de nombreux arbres feuillus (et l'énumération
va des espèces européennes comme Fagus silvatica et Quercus
pedunculata, aux espèces tropicales comme Mangletia glauca
et Erythrophleum fordii, en passant par celles du Pacifique ou de l'Australie)
se comportent un peu à la façon de Fagus silvatica : leurs
feuilles d'ombre sont caractérisées par une photosynthèse
modérée sous leur lumière de saturation spécifique,
et leur activité photosynthétique se stabilise sensiblement par
la suite. Mais leurs feuilles de lumière continuent à accroître
leur activité jusqu'à des éclairements de saturation bien
plus élevés.
En se plaçant du point de vue de la nutrition carbonée,
on enregistre donc une nette différence de comportement, vis-à-vis
de la lumière entre la majorité des arbres feuillus, et un certain
nombre de résineux.
Il est à noter du reste que, quelque soit la position
des appareils foliacés dans les cimes et le microclimat lumineux qui
y règne, on n'enregistre pratiquement pas de " photosynthèse
négative ", c'est-à-dire une perte de substances hydrocarbonées,
par la respiration, supérieure à la synthèse de ces mêmes
substances, réalisée par la photosynthèse. Tous les organes
chlorophylliens concourent donc, d'une façon très différente
évidemment, mais la plupart du temps positive à la nutrition carbonée
de l'arbre qui les supporte. Ceci sauf de nuit, bien entendu.
CONCLUSION
En somme, dans un certain nombre de cas (arbres résineux
notamment), les considérations qui précèdent atténuent
un peu l'importance que l'on pensait, au début, pouvoir attacher à
la considération des niveaux d'énergie dans les peuplements forestiers.
On retrouve, et l'on retrouvera sans doute de plus en plus, au fur et à
mesure que les études se développeront et se préciseront,
que l'arbre n'est pas uniquement une machine, aux réactions impérativement
dictées par les conditions physiques, mais également un être
vivant, qui, de lui-même, s'efforce de tirer le meilleur parti du microclimat
dans lequel il est établi.
Ses possibilités d'adaptation sont cependant assez
strictement limitées : une régénération de chêne
pédonculé, dont les feuillages ne reçoivent qu'un éclairement
énergétique de 150 à 200 kW / ha (*) est destinée
à disparaître rapidement - une plantation d'épicéa
maintenue sous un éclairement énergétique de 300 à
400 kW / ha (*) ne se développera que d'une façon très
lente, etc.
On peut donc, à titre de première approximation, continuer à élaborer les grands traits d'une analyse systématique de l'utilisation de l'énergie dans les peuplements forestiers, selon, par exemple, les types de raisonnement exposés aux paragraphes ci-dessus (et selon beaucoup d'autres, évidemment...). Mais il conviendra de ne jamais considérer ces positions comme définitives, et, au fur et à mesure que des relations plus précises (du genre de celles exposées au paragraphe IV ci-dessus) seront mises en évidence, il importera de corriger les données obtenues, de les nuancer, afin de se rapprocher, peu à peu, d'un mode de raisonnement plus conforme à la très complexe réalité.
(*) puissance moyenne ramenée à une durée d'ensoleillement de 12 heures par jour — en multipliant cette valeur par 12, on obtient une quantité d'énergie par hectare exprimée en kWh.
BIBLIOGRAPHIE
(1) ROUSSEL (L.). - Les radiations naturelles et la forêt.
Bulletin trimestriel de la Société forestière de Franche-Comté,
n° 2 et n° 4, 1966 ; n° 8, 1967.
(2) GESLIN (H.), BOUCHET (R.J.). - Climat et production
végétale : la bioclimatologie. In : L'Institut National de
la Recherche Agronomique. Edition du 20° anniversaire 1946-1966, 1966,
pp. 35-58 (Regards sur la France n° 32, sept. 1966).
(3) GALOUX (A.). - Approche énergétique
et cybernétique de la hêtraie. Lejeunia, no 23, 1963.
(4) PERRIN de BRICHAMBAUT (Ch.). - Rayonnement solaire
et échanges radiatifs naturels. - Paris, Gauthier-Villars, 1963.
(5) CHARTIER (Ch.). - Lumière, eau et production
de matière sèche du couvert végétal. Annales agronomiques,
vol. 18, n° 3, 1967, pp. 301-331.
(6) GRULOIS (J.). - Extinction du rayonnement global,
tropismes et paramètres foliaires. Bulletin de la Société
royale de botanique de Belgique, vol. 100, fasc. 2, 1967, pp. 315-334.
(7) ROUSSEL (L.). - Étude théorique élémentaire
de la trouée et de la bande. Bulletin trimestriel de la Société
forestière de Franche-Comté, n° 2, juin 1962, pp. 552-560.
(8) PISEK (A.), TRANQUILLINI (W.). - Assimilation und
Kohlenstoffhaushalt in der Krone von Fichten und Rotbuchenbäumen. Allgemeine
Botanische Zeitung, Band 141, 1954.
(9) PISEK (A.), WINKLER (E.). - Licht und Temperaturabhangigkelt
der C02. Assimilation von Fichte, Zirbe und Sonnenblume. Planta,
n° 53, 1959.
(10) LARCHER (W.). - The effect of environmental and physiological variables on the carbon dioxide gas exchange of trees. Photosynthética, vol. 3, n° 2 1969.