La plus importante réaction chimique de la terre
La photosynthèse (1)
par Louis Roussel

Photo Robbe.
Article publié en 1976 dans la Revue Bois et Forêts des Tropiques, n° 168, p19 - (version .pdf - 1,16 M)
|
La plus importante réaction chimique de la terre La photosynthèse (1) par Louis Roussel
|
|
| Le Pinus
merkusii est une essence de lumière totale. Sur cette photo, le
sujet surcimé dépérit et disparaîtra en quelques
années. Cambodge. Photo Robbe. |
SUMMARY
OUR PLANET'S MOST IMPORTANT CHEMICAL REACTION - PHOTOSYNTHESIS
After a brief reference to the importance of photosynthesis, the author reviews
the principal stages which led to modern ideas concerning the details of how
this phenomenon occurs.
He then examines the structures of the foliar mechanism and its reactions to variations in environmental factors. Passing from the leaf to the tree, simple examples are given of the determination of relative compensation lighting and relative saturation lighting for certain forest species in a natural environment or under cultivation. Lastly, the author shows how the increase in standing timber in general can be linked with the quantity or light energy absorbed by treetops.
Il est à peu près admis, actuellement, que les végétaux pourvus de chlorophylle élaborent, sur l'ensemble du Globe, grâce à la lumière naturelle, au dioxyde de carbone atmosphérique, à l'eau et à quelques substances minérales, d'énormes quantités de matières, d'un tonnage infiniment supérieur à celui produit par toutes les activités industrielles humaines. Cette réaction est, actuellement, présentée de la façon suivante, qui tient compte, comme il sera exposé plus loin, du fait que l'oxygène, finalement rejeté, provient de la décomposition de l'eau du sol :
![]()
À vrai dire, étant donné la variété considérable
des diverses formations végétales, et les grandes différences
qui existent entre leurs activités photosynthétiques respectives,
on a avancé, depuis une trentaine d'années, des chiffres assez
dissemblables. En 1945, aux U. S. A., l'un des biochimistes qui ont contribué
le plus efficacement à faire progresser les connaissances en ce domaine,
RABINOWITCH, estimait la fixation annuelle du carbone atmosphérique à
155 milliards de t, pour les océans, et à 19 milliards de t, pour
l'ensemble des terres émergées. D'où une synthèse
théorique de polyholosides (du type amidon, cellulose, lignine, etc...)
" de l'ordre" de 400 milliards de t par an, au total, mais avec
une supériorité très marquée pour la photosynthèse
des plantes marines. En 1964, KOBAK, excellent spécialiste de ces problèmes
en U. R. S. S., réduisait sensiblement ce chiffre à environ 100
milliards de t de matières sèches, synthétisées
chaque année, avec une supériorité très atténuée
pour les plantes marines.
Lors du Colloque organisé par l'UNESCO, en 1969, sur la productivité
des écosystèmes terrestres et marins, des conclusions différentes
ont été avancées, étayées, semble-t-il, par
des estimations bien plus nombreuses et bien plus précises que celles
qui avaient conduit aux chiffres précédents. Ce sont les éléments
les plus récents diffusés sur cet important problème :
pour l'ensemble du Globe, la photosynthèse produirait, chaque année,
environ 164 milliards de t de matières sèches, dont 55 pour les
océans, et 109 pour les terres émergées (y compris les
rivières, lacs, étangs, marais, etc... ). Les forêts tropicales
interviendraient pour 40 milliards de t par an, les forêts tempérées,
pour 23 milliards de t, les forêts boréales pour 10 milliards de
t, les savanes pour le même tonnage, et les broussailles désertiques
pour 1,3 milliard de t. L'accroissement, par hectare et par an, varierait ainsi
de 0,7 t, dans le cas de ces dernières formations, à 20 t, pour
les forêts tropicales humides (avec une " fourchette " de 10
à 50 t). Il s'agit, bien entendu, de la formation annuelle de la "
biomasse " (troncs, branches et racines, avec leurs écorces, feuillages,
semences, etc... ) ; la partie utilisable économiquement, selon les habitudes
commerciales actuelles, ne représente guère que le 1/3 ou le 1/4
des tonnages ci-dessus. Quant à la proportion, effectivement exploitée,
de cet accroissement, on sait qu'elle est encore très nettement inférieure.
En tout cas, ces chiffres récents mettent en évidence la relativement
faible productivité des formations végétales marines, sur
lesquelles on avait pourtant beaucoup compté (1,25 t par ha et par an,
exception faite des estuaires et des régions côtières),
et la grande supériorité des formations forestières, qui
réaliseraient, à elles seules, la moitié de la production
végétale de l'ensemble du Globe. On peut penser, bien entendu,
que ces chiffres ne sont pas définitifs.
Quelles sont les idées actuelles sur les mécanismes intimes de
ce phénomène primordial ? Comment est-on arrivé à
nos conceptions modernes ? C'est ce qui va être exposé ci-après.
On ne retiendra, du reste, que certaines des étapes, ou que quelques-uns
des éléments qui auront semblé essentiels à l'auteur
de cette analyse, mais qui, de toutes façons, ne représenteront
qu'une faible partie de ce qu'il aurait fallu dire sur ce très vaste
et très important sujet d'études et de réflexions. Au surplus,
on parlera surtout des espèces forestières de l'Europe et de l'Amérique
du Nord, qui sont étudiées depuis un certain nombre d'années
; l'extension de ces recherches aux espèces tropicales est bien plus
récente, et loin d'être achevée.
LES DÉCOUVERTES FONDAMENTALES
UNE LONGUE COHORTE DE CHERCHEURS
Les prêtres de l'ancienne Egypte, au Moyen Empire, enseignaient déjà
que c'était RA, le soleil, qui faisait croître les plantes et conditionnait
toute la vie ; ceci près de quatorze siècles avant J.-C. Mais,
un millénaire plus tard ARISTOTE, avec l'immense autorité qui
s'était attachée à son nom, imposa ses vues de philosophe-naturaliste
à de nombreuses générations de savants. Il affirmait, par
exemple, que les végétaux se développaient grâce
aux matières diverses digérées par le sol, devançant
ainsi les futurs tenants de la " théorie de l'Humus ". Les
feuilles vertes n'étaient que des ornements, ou, à la rigueur,
des sortes de pare-soleil, et, pendant deux mille ans, parce qu'ARISTOTE "
l'avait dit ", on ne chercha pas plus avant.
Et puis, brusquement, tout se mit à changer vers la fin du XVIIIe siècle ; en quelques décennies, des esprits perspicaces et observateurs : MALPIGHI, PRIESTLEY, LAVOISIER, INGEN HOUSE, SENEBIER, DE SAUSSURE... montrèrent que c'était la lumière solaire qui, grâce à l'eau et aux matières minérales du sol, ainsi qu'au gaz carbonique de l'air, provoquait, dans les feuilles des végétaux, la synthèse de toutes les substances qui leur permettaient de vivre, et de croître. Bien plus, ces mêmes végétaux rendaient l'air favorable à la vie des êtres animés, en y rejetant de l'oxygène (en le " dephlogistiquant ", pour reprendre l'expression des premiers savants de cette époque, d'intense remise en question des connaissances scientifiques). Tout paraissait alors assez net, et l'on pensa à ce moment qu'il n'y avait plus grand chose à découvrir de nouveau. Mais, d'éminents esprits restaient insatisfaits : LIEBIG, MAYER, BAEYER, SACHS, C. BERNARD, et toute une foule de chercheurs établirent, par la suite, que c'était la chlorophylle, à laquelle on avait peu prêté attention jusqu'alors, qui était l'agent principal de cette action, que l'on pouvait suspendre ses fonctions en anesthésiant les feuillages et que le volume de gaz carbonique absorbé était, d'abord voisin, puis égal, au volume de l'oxygène rejeté.
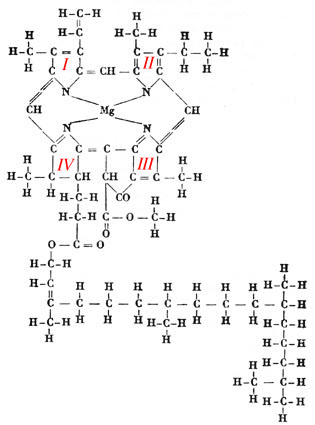
FIG. 1. - Formule développée
de la chlorophylle a, selon J. Carles. Remarquer le groupement caractéristique
(un atome de Mg entouré de 4 atomes de N, appartenant aux quatre noyaux
pyrroliques, numérotés de I à
IV).
Il y a à peu près un siècle, GRANDEAU, dans le cours d'Agriculture qu'il professait à l'Ecole Forestière de Nancy, soulignait l'importance " de cet acte fondamental de la vie des plantes ", accompli grâce à la chlorophylle (de composition encore inconnue en 1879), mais reconnaissait que l'on était dans la plus complète ignorance des détails des mécanismes de ce processus. Il soutenait fermement, du reste, les vues de LIEBIG, contre les idées de THAER, éminent porte-parole des tenants de la théorie de l'humus.
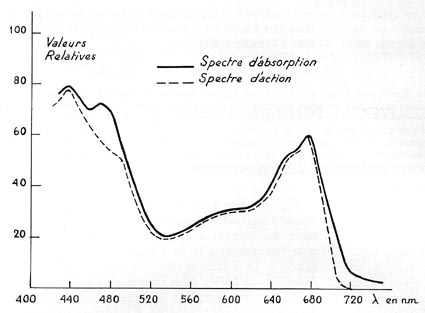
FIG. 2. - Le spectre d'absorption de la feuille,
soumise à des radiations de différentes longueurs d'onde (pointes
au voisinage de 440 et 680 nm), concorde assez bien avec le spectre d'action
des mêmes radiations (caractérisé par les quantités
de CO2 fixées, par une feuille identique, et dans des
conditions énergétiques égales). Reproduit par A. Moyse
(nm = 10-9 m).
Un demi-siècle plus tard, vers 1925, on était un peu plus avancé
: BLACKMANN avait montré qu'à côté des réactions
provoquées indiscutablement par la lumière, d'autres se poursuivaient
très bien à l'obscurité. WILLSTAETTER & STOLL, et leurs
émules, avaient proposé des formules chimiques pour les deux formes
de chlorophylles découvertes, a et b. WURMSER avait commencé à
déterminer les spectres d'absorption de ces chlorophylles en solution
dans l'acétone et dans l'éther, ou bien telles qu'elles se présentaient
dans des algues marines. Des expérimentateurs ingénieux, comme
TIMIRIAZEFF, ENGELMAN, DANGEARD, et autres, montraient que c'étaient
les radiations absorbées dans le bleu, et dans le rouge, qui étaient
de beaucoup les plus actives pour stimuler ce que l'on appelait : l'assimilation
chlorophyllienne. Mais, les mécanismes intimes étaient encore
inconnus. Une théorie avait alors un certain succès : la synthèse
des glucides (du type C6H12O6, comme le glucose)
résultait de la jonction de 6 molécules d'aldéhyde formique
(CH2O), formé, lui-même, d'une façon très
simple : CO2 + H2O = CH2O +O2. Par
perte d'une molécule d'eau, le glucose fournissait l'élément
de base : C6H10O5, dont la réunion en
longue chaîne donnait l'amidon, la cellulose, et autres polyholosides.
Cependant la toxicité, bien connue, de l'aldéhyde formique, rendait
cette hypothèse difficile à accepter par de nombreux botanistes
; c'est pour cette raison que MARQUENNE, à cette même époque,
proposait d'admettre que la chlorophylle entrait en combinaison temporaire avec
le gaz carbonique, ce qui écartait l'objection précédente.
Certains chercheurs, aux Etats-Unis, commençaient à employer
des méthodes originales : EMERSON, par exemple, qui soumettait les végétaux
à de brefs éclairs de lumière, afin de tenter de déterminer
l'importance relative des réactions lumineuses, et obscures, dans le
déroulement de ce que l'on commençait à appeler "
la photosynthèse". En effet, jusqu'alors, on étudiait plutôt
" l'assimilation chlorophyllienne ", la " décomposition
du gaz carbonique ", ou " l'assimilation du carbone ".
LA FÉCONDE PÉRIODE AMÉRICAINE
Vers le milieu du XXe siècle, au moment où se préparait,
puis se déclenchait la seconde guerre mondiale, dans leurs laboratoires,
aux Etats-Unis surtout, les chercheurs poursuivaient l'étude de la photosynthèse,
et les progrès considérables qui ont été réalisés
récemment sont dus, pour une bonne part, à l'emploi de techniques
très nouvelles :
- l'utilisation des "isotopes " (éléments chimiquement
identiques, mais dissemblables par leurs propriétés radioactives,
ou leurs masses atomiques) ;
- la " chromatographie ", dont on rappellera seulement qu'elle
utilise la vitesse différente de déplacement sur un papier filtre
de substances organiques complexes, selon leur nature, et selon le solvant employé.
C'est ainsi que RUBEN, en 1941, en utilisant un isotope lourd (mais non radioactif)
de l'oxygène : 18O, montra, d'abord, que c'était l'eau,
et non le gaz carbonique, qui fournissait ce gaz, indispensable à tous
les êtres vivants.
La très complexe incorporation du gaz carbonique à la substance
vivante de la plante fut suivie surtout en utilisant des isotopes radioactifs
du carbone , d'abord, le 11C, d'un emploi difficile, car sa période
(ou temps pendant lequel sa radioactivité est réduite de moitié)
est brève et voisine de 20 mn. Puis, le 14C, bien plus stable,
puisque sa période dépasse 5.700 ans. Les divers éléments,
successivement synthétisés par les plantes (en général
des algues marines, qui furent les plus utilisées), à qui l'on
a fourni du dioxyde de carbone marqué au 14C, sont dissociés
par la méthode de la chromatographie. Puis, le papier filtre qui les
supporte est placé, à l'obscurité, contre une émulsion
sensible aux rayons X (émis par le 14C), et les éléments
dont l'on recherche la nature et la présence s'inscrivent d'eux mêmes,
par "autoradiographie", sur l'émulsion. Ces procédés
furent à l'origine d'une période d'intense activité pour
les chercheurs des Etats-Unis : après RUBEN, HASSID et KAMEN, utilisant
le 11C, ce fut l'équipe dirigée par CALVIN, avec BASSHAM,
BENSON, MASSINI, WILSON notamment, qui grâce à l'emploi du 14C,
contribua le plus efficacement à élucider, tout au moins en grande
partie, le mécanisme intime de la photosynthèse. Le schéma
qui résume leurs travaux est appelé, en général,
" Cycle de CALVIN " (ou " Cycle de CALVIN-BASSHAM ")
du nom de l'animateur de cette équipe qui reçut, en 1961, le prix
Nobel de chimie.
Voici, du reste, comment CALVIN et BASSHAM analysent sommairement le mécanisme
de la photosynthèse :
" L'eau est d'abord décomposée, par une réaction
photolytique, en donnant un réducteur et un composé intermédiaire
instable, qui libère l'oxygène ; ensuite, le réducteur
sert à réduire le gaz carbonique, par une, ou plusieurs réactions
obscures. " D'une façon assez laconique, ces auteurs indiquent que
le réducteur est [H], et le composé qui libère l'oxygène
[OH], selon le processus
2 [OH] --> 1/2 O2 + H2O
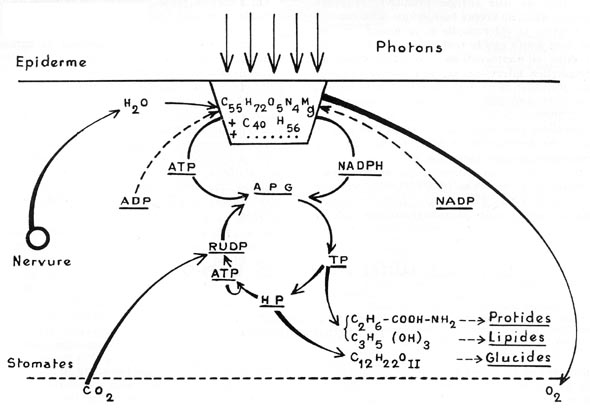
FIG. 3. - Schéma très simplifié
du " Cycle de CALVIN ".
L'eau du sol (H2O) arrive, par les nervures, dans les
cellules à chloroplastes (C55H72O5N4Mg
= chlorophylle a + C40H56 = carotène
+ ...), soumises à un flux de photons. L'oxygène est rejeté
par les stomates (O2) : l'adénosine diphosphate (ADP)
est transformée en adénosine triphosphate (ATP), et
le nicotinamide adénine dinucléotide phosphate (NADP) est
réduit en NADPH ; ATP et NADPH sont enrichis en énergie
chimique.
En présence du ribulose diphosphate (RuDP), carbonaté
par le gaz carbonique de l'air (CO2), pénétrant
par les stomates, se forme l'acide phosphoglycérique à 3 atomes
de carbone (APG), puis des sucres trioses phosphates (TP), puis des sucres
hexoses phosphates (HP) qui, d'une part, donnent du saccharose (Cl2H22O11),
base de la synthèse des glucides, d'autre part, avec les TP,
et grâce à un nouvel apport d'ATP, reforment du RuDP,
qui rentre dans le cycle. En outre, les TP donnent des acides aminés
(du type alanine (C2H6-COOH-NH2)), base
de la synthèse des protides, et également du glycérol (C3H5
(OH)3), base de la synthèse des lipides.
REMARQUE: Pour rendre le schéma plus concret, il a été
situé dans une feuille coupée transversalement ; la photolyse
de l'eau a lieu, très probablement, dans les cellules palissadiques situées
dans la partie supérieure (la mieux éclairée) de la feuille.
Mais, pour le surplus du " Cycle ", il ne faut absolument pas rechercher
une quelconque localisation des diverses réactions.
En ce qui concerne l'énergie nécessaire à ces opérations,
les photons (minuscules parcelles d'énergie qui, en nombre extrêmement
grand, constituent le flux lumineux qui atteint les feuillages des végétaux),
" excitent " des groupes de molécules de chlorophylle (associées
en " antennes collectrices " et en " centres réactionnels
"), en déplaçant certains de leurs électrons sur des
orbites plus externes. Quand ces électrons reviennent sur les orbites
primitives, ils libèrent une énergie chimique qui va se concentrer,
temporairement, dans des sortes " d'accumulateurs ", molécules
organiques complexes, caractérisées par la présence d'atomes
de phosphore, et dont les formules détaillées ne seront pas données
ici :
- l'ADP (adénosine diphosphate) qui se transforme en ATP (adénosine
triphosphate) ;
- le NADP (nicotinamide adénine dinucléotide phosphate) qui se
transforme en NADP H2 (forme réduite).
Tout n'est pas encore clair, dans ces problèmes de transfert d'énergie
; on a dit que la chlorophylle absorbait spécialement deux groupes de
radiations : les radiations bleues, dont la longueur d'onde est voisine de 440
nm, et les radiations rouges, dont la longueur d'onde est voisine de 680 nm.
Or, il semble que ce sont surtout ces dernières radiations qui joueraient
un rôle important dans le mécanisme des transferts d'énergie
: les premières interviendraient aussi, mais après avoir perdu,
sous forme calorifique une partie de leur énergie primitive, et étant
ramenées, ainsi, au niveau énergétique des secondes. Par
ailleurs, la chlorophylle a, la mieux connue, n'est sans doute pas le seul pigment
qui joue un rôle dans la photosynthèse : d'autres types de chlorophylles
interviennent, et aussi les caroténoïdes (carotène et xantophylle).
De toutes façons, on accorde actuellement une grande importance à
deux groupes de pigments, le P680, et le P700, qui
absorbent les radiations de 680 et de 700 nm, et constituent, dans les études
théoriques de transferts d'énergie, les photosystèmes I
et II.
Dans le " Cycle de CALVIN ", qui ne comprend pas moins de 14
réactions (nécessitant la présence de plus de 10 sortes
d'enzymes) le premier produit synthétisé est l'acide phosphoglycérique
(APG), a 3 atomes de carbone ; d'où le nom de végétaux
en C3 donné aux plantes, extrêmement nombreuses, qui
suivent ce cycle, et qui intéresse tout spécialement les sylviculteurs.
Assez récemment, cependant, KORTSCHAK, HARRT et BURR, puis HATCH et
SLACK, faisaient connaître (1965-1970) que certaines graminées
tropicales (le maïs et le sorgho, par exemple) utilisaient une voie différente
: le premier produit élaboré était un oxaloacétate
(comprenant 4 atomes de carbone). Après quelques hésitations et
quelques vérifications, le cycle en C4 a été
admis ; les végétaux qui s'y conforment représenteraient
environ 180 espèces appartenant pour une bonne part à la famille
des Graminées. À première vue, ce type de plante n'intéresse
pas, directement, les forestiers.
Enfin, certaines plantes grasses (des Crassulacées) fixent, de nuit,
le dioxyde de carbone atmosphérique, pour l'utiliser, pendant le jour
à leur photosynthèse.
Si l'on compare l'activité photosynthétique des plantes en C3
avec celle des plantes en C4, on peut remarquer que les premières
s'arrêtent d'assimiler le carbone, quand la teneur de l'atmosphère,
en dioxyde de carbone (normalement voisine de 0,03 %), descend au-dessous de
0,005 %, alors que les secondes continuent à fonctionner jusqu'à
une teneur de l'ordre de 0,001 %. Par ailleurs, les plantes en C3
n'assimilent, dans les meilleures conditions, que la moitié environ du
carbone retenu, dans les mêmes conditions, par les plantes en C4.
On a surtout parlé, jusqu'ici, de la seule photosynthèse, au
sens strict du terme, c'est-à-dire de l'acte par lequel le végétal
se nourrit, comme il a été dit, grâce à la lumière,
au dioxyde de carbone atmosphérique, à l'eau et à quelques
matières minérales du sol. Or, actuellement, on étend parfois
cette étude à la respiration, obscure, ou bien stimulée
également par la lumière, et que l'on nomme alors " photorespiration
" ; il s'agit, dans ce cas, des fonctions par lesquelles le végétal
réutilise les substances provenant de la photosynthèse qui, oxydées
grâce à l'oxygène atmosphérique, lui permettront
de satisfaire ses besoins énergétiques. Ce genre d'étude,
très intéressant en lui-même, sort nettement du cadre, obligatoirement,
restreint, de cette analyse.
LA FEUILLE, MAITRE D'OEUVRE DE LA PHOTOSYNTHÈSE
STRUCTURE DE LA MACHINERIE VÉGÉTALE
Bien que l'on ait décelé une activité photosynthétique
dans toutes les parties vertes des végétaux (jeunes tiges comprises),
c'est, évidemment, la feuille qui est l'organe de beaucoup le plus important
pour l'accomplissement de cette fonction essentielle. On ne s'étendra
pas sur leur description détaillée : qu'il s'agisse des aiguilles
des résineux, ou bien parfois des écailles, soudées aux
rameaux des Cyprès, ou, au contraire, des feuilles plus ou moins largement
étalées des arbres dits feuillus, elles ont toutes le même
rôle, et l'on y discerne toujours un certain nombre de régions
principales : une assise de cellules externes, l'épiderme, au-dessous,
des cellules renfermant la chlorophylle, souvent de forme allongée, ou
palissadique, mais aussi sans grande différenciation parfois avec la
forme de leurs voisines. Enfin des cellules collectrices qui recueillent les
produits élaborés dans les cellules à chlorophylle et les
conduisent vers les nervures qui évacuent lesdits produits vers l'axe
central. De minuscules ouvertures, ou stomates, localisées, en général,
sur la face inférieure des feuilles des arbres feuillus, ou bien sur
l'ensemble du pourtour des aiguilles des arbres résineux, permettent
au dioxyde de carbone de pénétrer dans la machinerie végétale,
à l'oxygène et à l'eau de transpiration d'en sortir. À
côté des stomates, cependant, il est de nombreux cas où
les mouvements de l'eau et des gaz se font, d'une façon le plus souvent
réduite, au travers de l'épiderme lui-même, recouvert d'une
cuticule de protection plus ou moins épaisse, et plus ou moins étanche.
La structure des feuilles dépend, assez fréquemment,
des conditions microclimatiques sous lesquelles elles se sont développées
; pour les sujets du genre Quercus, des études anatomiques nombreuses
ont été effectuées, en particulier à l'Institut
Botanique de Besançon (TRONCHET, GRANDGIRARD, GIBOUDEAU, GOGUELY), en
liaison avec le Service forestier local. Par exemple, pour Q. pedunculata,
l'épaisseur de la feuille augmente régulièrement, de
100 µ environ, quand le sujet s'est développé sous un couvert
assez dense, jusqu'à 170 µ environ, pour le sujet développé
en plein découvert ; la cuticule supérieure, qui protège
la feuille contre l'excès de radiations, présente, dans les mêmes
conditions, un triplement d'épaisseur (2,4 µ à 6,4 µ). Les cellules
palissadiques, renfermant la majorité de la chlorophylle, doublent de
longueur (24,5 µ à 54 µ) ; le tissu lacuneux, où circulent les
substances diverses, gazeuses ou liquides, voit son épaisseur passer
de 40 à 74 µ. Le nombre des stomates, par lesquels se font les échanges
avec le milieu extérieur, augmente sensiblement, quand le sujet passe
de l'ombre assez dense (200 stomates au mm2) à la pleine lumière
(400 stomates au mm2) . Grâce à la technique délicate
des coupes faites parallèlement à la surface des feuilles, on
a pu montrer que le volume des cellules palissadiques (où se déroule
l'essentiel des réactions de la photosynthèse), est triplé
quand le sujet passe de l'ombre à la pleine lumière.
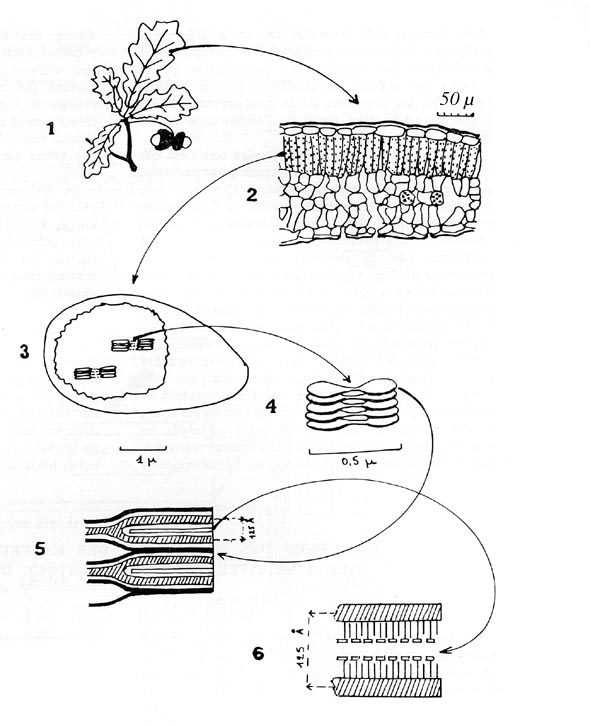
FIG. 4. - Anatomie simplifiée de la feuille de
chêne pédonculé.
1. - Bouquet de feuilles de chêne pédonculé (Quercus
pedunculata).
2. - Coupe transversale d'une feuille développée à la
lumière. De haut en bas : cuticule et épiderme, cellules palissadiques
allongées renfermant de nombreux chloroplastes, cellules collectrices,
chambres sous-stomatiques et stomates, dans l'épiderme inférieur.
3. - Un chloroplaste partiellement ouvert, laissant voir les grana, baignant
dans le stroma filamenteux.
4. - Une pile de grana, sortes de disques unis par des filaments, et renfermant
la chlorophylle.
5. - Coupe perpendiculaire à la surface des grana, laissant voir leur
structure intime.
6. - Même coupe agrandie : la partie hachurée représente
une association de molécules de protéine, les petits traits verticaux,
des molécules de lipides, et les petits traits verticaux surmontés
d'un petit plateau, des groupes de molécules de chlorophylle réunies
en antennes collectrices de lumière et en centres réactionnels.
Echelles des dessins : µ = 10-6 m ; Å = 10-10 m (d'après des dessins de A. GRANDGIRARD, J. DE ROSNAY, J. A. BASSHAM et M. CALVIN).
Pour les résineux, diverses études ont déjà été
effectuées ; dans le cas de Picea excelsa, par exemple, si l'aiguille
du sujet développé à l'ombre dense (environ 3 % d'éclairement
relatif) a une section peu inférieure à celle du sujet développé
en pleine lumière (les 5/6 environ de celle-ci), la cuticule est nettement
plus développée chez ce dernier sujet (épaisseur de 3 µ,
contre 1 µ environ pour le sujet d'ombre). Les cellules de l'épiderme
ont des membranes un peu plus minces chez le sujet d'ombre ; on ne distingue
pas, chez cette espèce, de parenchyme palissadique chlorophyllien individualisé,
comme chez les feuillus, mais plusieurs couches concentriques de parenchyme
lacuneux, qui renferment la chlorophylle. La densité des stomates varie
assez peu et reste " de l'ordre " de 50 au mm2, chiffre
nettement inférieur à celui observé, en général,
chez les arbres feuillus. AUSSENAC a trouvé, sur divers sujets des genres
Abies, Pseudotsuga et Picea, des différences plus marquées
entre les aiguilles d'ombre et les aiguilles de lumière, dans le sens
indiqué plus haut pour les arbres feuillus. La surface des stomates,
élément important, a été étudiée à
Innsbruck. Ce genre de recherche est relativement encore peu développé,
bien qu'il constitue, cependant, une approche pratique des problèmes
de la photosynthèse des espèces forestières.
La chlorophylle est localisée dans les chloroplastes, sortes de corpuscules
en général lenticulaires, répartis dans le cytoplasme des
cellules palissadiques, chez les arbres feuillus, comme les chênes, ou
bien dans le parenchyme lacuneux, comme chez de nombreux résineux, parfois
aussi autour des vaisseaux externes. Selon BOURDU leur nombre est très
variable (de 5 à 500 par exemple, dans une seule cellule selon les espèces)
; leur volume est également variable, de 30 à 300 µ3).
Chaque chloroplaste est constitué d'une double membrane périphérique,
entourant une sorte de gelée, le stroma, dans laquelle sont placés
des empilements de disques accolés, les grana. C'est dans ces grana que
sont disposées, en associations distinctes (antennes collectrices et
centres réactionnels), les molécules de chlorophylle.
INFLUENCE DES VARIATIONS DES FACTEURS DU MILIEU SUR L'ACTIVITÉ PHOTOSYNTHÉTIQUE
DES FEUILLES
La façon dont les feuilles réagissent, en ce qui concerne leur
activité photosynthétique, est étudiée, en général,
dans des appareils à absorption de rayonnement infrarouge (URAS dans
les pays de langue germanique et IRGA, dans ceux de langue anglaise). Bien entendu,
on peut faire intervenir de très nombreux facteurs, soit simultanément,
soit en séquence, et se placer, parfois, dans des conditions que les
végétaux ne rencontrent pas, pratiquement, dans le milieu naturel.
C'est le cas, par exemple, des variations artificielles dans la teneur, en dioxyde
de carbone, de l'air que l'on envoie dans ces appareils. On peut, par exemple,
montrer facilement que la plupart des arbres, résineux et feuillus, habitués
à une atmosphère contenant, en moyenne, 0,03 % de dioxyde de carbone,
assimilent davantage quand cette teneur augmente, jusqu'à 0,3 % environ.
Ceci, sans dommage apparent pour les végétaux en expérience.
Si l'on augmente encore la teneur de l'air en dioxyde de carbone, on obtient
souvent une majoration de l'assimilation carbonée, mais, cette fois,
avec quelques signes de fatigue, des nécroses, etc... Cependant, en restant
dans des limites assez strictes, on doit pouvoir améliorer nettement
la croissance des jeunes sujets en expérience, et ainsi, gagner un ou
deux ans dans l'obtention de plants utilisables pour les boisements. Mais, c'est
surtout l'influence de la lumière et de la température, sur l'activité
photosynthétique des feuilles et des aiguilles, qui a été
le mieux analysée, tout spécialement, d'abord, à l'Institut
Botanique d'Innsbruck et au Phytotron du Patscherkofel, en Autriche. Sous la
direction du Professeur PISEK, des chercheurs perspicaces comme TRANQUILLINI,
WINKLER, LARCHER et leurs collaborateurs ont accumulé les observations,
et ce qui est trouvé actuellement ne fait souvent que confirmer des résultats
publiés dès les années 1950.
Les observations ont porté sur la photosynthèse nette, telle qu'elle est analysée dans les appareils mentionnés plus haut : les chiffres correspondent aux quantités de carbone effectivement fixé par la feuille. Pour les études de physiologie végétale plus développées, on recherche la photosynthèse brute, en ajoutant aux chiffres obtenus ci-dessus, la respiration obscure (nocturne, dans le milieu naturel) et la photorespiration, plus délicate à déterminer, qui se produit pendant le jour.
))
FIG. 5. - L'URAS (ou IRGA), dont l'emploi se généralise,
est un appareil très important pour étudier, du point de vue pratique,
les réactions des feuillages aux variations de la lumière et de
la température, principalement. Les rameaux à observer, alimentés
en eau pendant le temps de l'expérience, sont placés dans les
cuvettes transparentes (photosynthèse nette), ou opaques (respiration
obscure). L'éclairement vient de la partie supérieure et peut
atteindre 50.000 lux ; la lumière est de composition voisine de celle
de la lumière naturelle. La température des chambres ou cuvettes
est réglée grâce à un système de circulation
d'eau, avec thermorégulation. L'air extérieur entre par la partie
gauche (pompes) et circule autour des rameaux, puis, plus ou moins appauvri
en CO2 (dans les cuvettes transparentes), ou enrichi en CO2
(dans les chambres opaques), passe dans un analyseur à rayons infrarouges
(partie inférieure du dessin), qui constitue l'URAS proprement dit. Les
indications relatives à l'enrichissement, ou à l'appauvrissement
de l'air en CO2 sont inscrites sur des graphiques (bas du dessin,
à gauche). La précision admise est de l'ordre de ± 3 %.

Khaya ivorensis. Plantation de Ngolon de 6 ans
bien éclairée par la route. Cameroun, 1957.
Photo Sarlin.
Le premier élément qui ressort de ces observations est que la
photosynthèse nette la plus élevée est obtenue,
sous chaque éclairement, pour une température des feuilles (ou
des aiguilles) bien déterminée, et qui augmente légèrement
quand les éclairements s'accroissent. La température trop basse
ne permet pas aux cellules à chlorophylle de fonctionner dans de bonnes
conditions ; la température trop élevée accentue les processus
respiratoires, qui réutilisent partiellement, et immédiatement,
les substances élaborées par la photosynthèse. Le végétal,
grâce à la transpiration (voir Bois et Forêts des Tropiques,
n° 146) peut lutter contre un échauffement excessif de ses feuilles
(ou de ses aiguilles), et, dans les meilleures conditions, se rapprocher des
conditions optimales de température, propres du reste à chaque
espèce, et à chaque éclairement.
Le second élément, qui intéresse plus spécialement les sylviculteurs, est que, dans une fourchette de températures, pas trop éloignées de l'optimum défini plus haut, l'intensité de la lumière règle, d'une façon très efficace, la marche de la photosynthèse ; mais cette influence favorable est plus marquée aux éclairements faibles et moyens, qu'aux éclairements élevés. C'est ce que l'on exprime en disant que " le coefficient d'utilisation de la lumière diminue quand l'intensité de l'éclairement s'accroît ", et l'on arrive, plus ou moins tôt, à un " palier de saturation ". Par ailleurs, on a décrit précédemment les modifications de structure que subissent les feuillages, selon qu'ils se seront développés sous des couverts plus ou moins denses, dans des trouées, ou bien, en plein éclairement ; or, l'activité photosynthétique de ces feuillages n'est pas la même, en conditions microclimatiques égales, les feuilles d'ombre ayant, en général, une faculté d'assimilation du carbone plus élevée, aux faibles éclairements, que les feuilles de lumière.
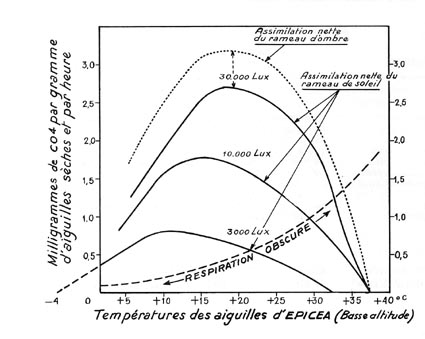
FIG. 6. - Courbes obtenues grâce à un appareil URAS, pour des rameaux d'épicéa commun (Picea excelsa), développé autour de 600 m d'altitude. La photosynthèse nette, par unité de poids, augmente avec l'intensité de l'éclairement, mais, dans chaque cas, le maximum n'est atteint qu'au voisinage d'une température des aiguilles bien déterminée (à quelques degrés près, évidemment). Même sous un éclairement assez élevé (30.000 lux), le rameau d'ombre assimile davantage que le rameau de lumière. Observations faites à Innsbruck par Pisek et Winkler.
La courbe " respiration obscure ", obtenue dans les chambres opaques, ne représente que la respiration nocturne ; la respiration diurne, ou photorespiration, est plus élevée.
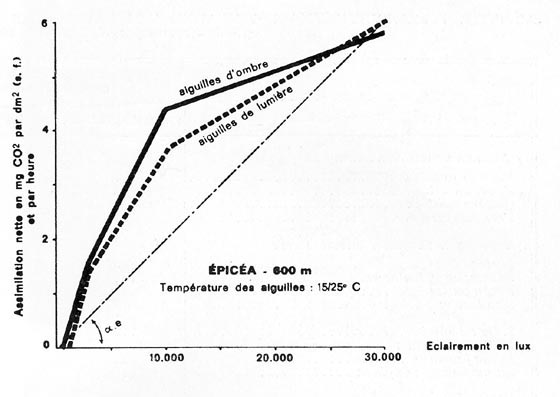
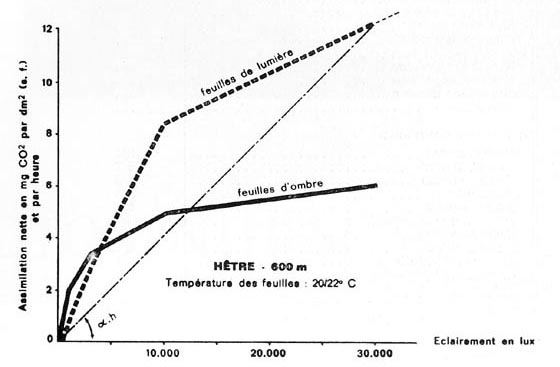
FIG. 7. - Courbes obtenues grâce à un appareil URAS, pour des rameaux de Picea excelsa, et de Fagus silvatica, se développant vers 600 m d'altitude. Influence de l'intensité de l'éclairement sur l'activité photosynthétique nette des aiguilles et des feuilles, dans une fourchette de températures normales en été.
Pour les organes foliacés d'ombre, et de lumière, le " coefficient d'utilisation de la lumière ", qui peut être matérialisé par tg alpha.e, et tg alpha.h., diminue constamment quand l'éclairement s'accroît. Réserves d'usage en ce qui concerne les chiffres des résineux, par unité de surface, probablement ici un peu sous-estimés.
D'après PISEK et WINKLER pour Picea, et LARCHER et RETTER, pour
Fagus.
Il est à noter aussi que l'on peut rencontrer, dans les mêmes
stations, des espèces à feuillages caducs, et d'autres à
feuillages persistants. Dans les régions où ces espèces
ont pu être étudiées de façon précise, on
observe que l'activité photosynthétique est plus élevée,
pour les premières, que pour les secondes. Il semble que les végétaux,
dont les feuillages ne " travaillent " pas très longtemps
dans une année, mettent, en quelque sorte, les bouchées doubles,
de façon a ne pas être trop distancés par leurs concurrents
immédiats (exemples : Larix europea en présence de Picea
excelsa, et Quercus pubescens en présence de Quercus ilex
selon les travaux de LARCHER). Pour préciser un peu ce qui vient
d'être dit au sujet de l'activité photosynthétique des feuilles
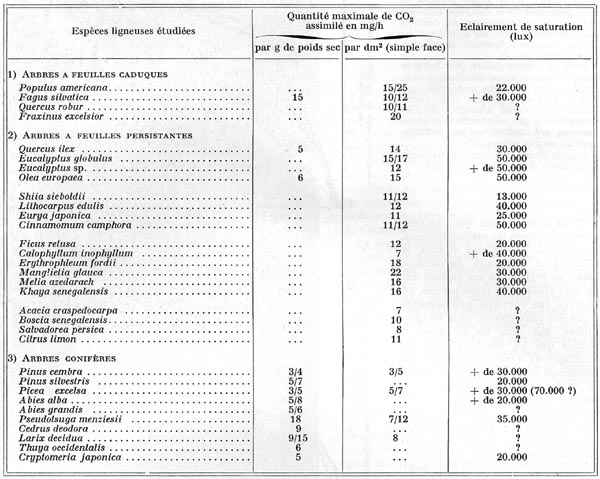
et des aiguilles, on donnera ci-après quelques chiffres, exprimés
en milligrammes de dioxyde de carbone assimilé par heure, pour un gramme
d'aiguilles ou de feuilles sèches, ou bien par décimètre
carré de feuillage. Une remarque importante est à faire sur ce
dernier point : pour des arbres feuillus, dont les feuilles sont très
minces, par rapport à leur surface, il n'y a aucune difficulté
à mesurer celle-ci : c'est l'élément " simple
face " qui figure dans de nombreux tableaux relatifs à la photosynthèse
des feuillages. Par contre, pour les aiguilles de résineux, comme leur
section va de l'ellipse plus ou moins allongée, au losange, ou au cercle,
on a dû adopter comme convention de considérer, comme simple face,
la moitié de leur surface totale. Or, du point de vue énergétique,
la surface réelle qui entre en ligne de compte est celle qui intercepte,
effectivement, le rayonnement naturel ; suivant donc la forme des aiguilles,
on attribue fictivement, à une surface conventionnelle, une activité
photosynthétique réalisée par une surface réelle,
en général inférieure. Il arrive donc assez souvent que
l'activité photosynthétique des aiguilles, formulée par
unité de surface conventionnelle, soit plus faible que leur activité
réelle, par unité de surface éclairée.
REMARQUES: On trouve, dans les tableaux de W. LARCHER, de multiples
autres indications, celles des " points de compensation ", par exemple,
c'est-à-dire des éclairements les plus bas (en général
de quelques centaines de lux) à partir desquels la plante fixe plus de
carbone, par la photosynthèse, qu'elle n'en perd par la respiration.
Au-dessous, la plante meurt rapidement par inanition énergétique.
Bien d'autres végétaux ligneux sont cités. D'une façon
générale, on ne relève pas une différence très
marquée entre la photosynthèse maximale des arbres des régions
tempérées, et celle des arbres des régions tropicales ou
équatoriales. Il faut aussi noter que les méthodes de travail
ne sont pas encore entièrement uniformisées, d'où les nombreux
vides qui existent dans ces tableaux.
ACTIVITÉ PHOTOSYNTHÉTIQUE COMPARÉE
DES FEUILLAGES DE DIVERSES ESPÈCES LIGNEUSES
(Chiffres réunis par W. LARCHER, 1969)