La plus importante réaction chimique de la terre
La photosynthèse (2)
par Louis Roussel

Photo Taylor.
Article publié en 1976 dans la Revue Bois et Forêts des Tropiques, n° 169, p51 - (version .pdf - 885 k)
|
La plus importante réaction chimique de la terre La photosynthèse (2) par Louis Roussel
|
|
| Régénération naturelle
de Khaya ivorensis et Entandrophragma utile au Ghana. Photo Taylor. |
PHOTOSYNTHESE ET CROISSANCES DES ARBRES
ET DES PEUPLEMENTS FORESTIERS
Il est bien évident que la croissance des arbres et des peuplements
forestiers a, comme support indispensable, leur nutrition en général,
c'est-à-dire, en fait, leur activité photosynthétique.
Mais, il n'y a pas, dans de nombreux cas, de concordance absolue, dans le
temps, entre cette activité photosynthétique, et la
croissance en longueur, en diamètre, en volume ou en poids des sujets
observés ; ceci tient au fait que les phénomènes de la
croissance sont liés, comme on l'a expliqué précédemment
(Bois et Forêts des Tropiques, no, 149 et 158) à divers
éléments, soit d'origine interne, comme certains rythmes biologiques,
soit d'origine externe, comme les variations de certains facteurs du milieu,
qui diffèrent parfois de ceux qui conditionnent la photosynthèse,
Le fait est particulièrement évident pour les résineux
des régions tempérées, par exemple, qui conservent des
aiguilles pendant toute l'année, et qui auront, pendant 8 à 10
mois, par exemple, une activité photosynthétique indéniable,
alors que leur croissance peut ne se manifester que pendant 3 ou 4 mois seulement.
Il semble que, dans les régions équatoriales, il existe bien une
certaine périodicité annuelle dans la croissance en longueur et
en diamètre de nombreuses espèces forestières (périodicité
liée davantage aux saisons sèches et humides, qu'au photopériodisme,
peu marqué) ; mais ces rythmes sont difficilement décelables.
La période de croissance, dans ces régions, est certainement plus
longue que dans les régions plus nordiques, et le bois d'apparence plus
homogène (CHEVALIER et NORMAND).
Que deviennent donc les matières élaborées, et non immédiatement
utilisées à la croissance ? une partie non négligeable
sert à entretenir les besoins énergétiques des arbres,
et ces processus se traduisent par les phénomènes respiratoires
(respiration diurne et nocturne des feuilles, respiration permanente du surplus
du corps de l'arbre, proportionnelle à sa surface extérieure,
laquelle ne représente, très souvent, que 15 à 20 % de
la surface des feuillages). Une autre partie des substances synthétisées
est évacuée des feuilles et mise en réserve ; leur migration
a été suivie par divers chercheurs : BONNEMAIN, KURSANOV, EVANS,
notamment, grâce en particulier au 14C dont il a été
parlé plus haut. Des feuilles, ces substances passent dans les "
tubes criblés " verticaux, élément essentiel du "
liber ", situés, grossièrement, entre le bois et l'écorce.
La vitesse de transfert longitudinal est élevée, et de l'ordre
de 1 m par heure (BONNEMAIN). En outre, il existe des transferts transversaux,
par les rayons libériens et ligneux (files de cellules disposées
horizontalement, et se prolongeant du liber jusqu'au cylindre central) ; mais
ces transferts sont plus lents (1 cm par heure), et ils se terminent par une
mise en réserve, dans les cellules ligneuses souvent : les
glucides, par exemple, transportés sous forme de saccharose soluble,
sont déposés sous forme d'amidon insoluble. Les oléorésines
des arbres résineux correspondent également à ce type de
mise en réserve (DAVID). Ils sont transformés, à nouveau,
en produits solubles, au moment de la reprise annuelle de la croissance, et
ils servent, avec des lipides et des protides, également stockés,
à la formation des nouvelles pousses feuillées.
D'un autre côté, les forestiers ont tendance à ne considérer
que la formation du bois du tronc, économiquement la plus intéressante
partie de l'arbre ; mais, outre le fût proprement dit, cet arbre doit
faire des feuilles, des branches, des racines, des semences aussi, qui tombent
périodiquement au sol et cette biomasse, toute entière, ne peut
s'élaborer que grâce à la photosynthèse.
Par ailleurs, pour que la photosynthèse elle-même puisse s'accomplir
dans de bonnes conditions, il faut que l'ensemble des facteurs du milieu
soit présent dans des proportions convenables, et si, un seul d'entre
eux est déficient, la photosynthèse sera réglée
par ce facteur. C'est ce que l'on appelle habituellement : " la
loi du minimum ". Ceci est particulièrement évident pour
les deux éléments climatiques qui paraissent les plus importants
: l'énergie lumineuse et l'eau du sol, et il n'est pas besoin d'insister
sur le fait qu'il sera tout aussi inefficace d'arroser abondamment un jeune
arbre forestier privé de lumière, que de le suréclairer,
s'il se développe sur un sol absolument sec... Ceci est vrai, aussi,
pour certains facteurs édaphiques : certes, la matière organique
totale d'une forêt comprend à peu près 98,5 % de carbone,
d'hydrogène et d'oxygène, éléments qui proviennent
de l'air et de l'eau du sol, mais le surplus, soit 1,50 %, peut parfois se révéler
important. Dans la chênaie-hêtraie à charmes de Virelles
Blaimont, en Belgique, DUVIGNEAUD et ses collaborateurs ont trouvé, sur
une masse totale de 156.000 kg par ha, 1.248 kg de calcium (0,80 %) 533 kg d'azote
(0,34 %) 342 kg de potassium (0,22 %) 102 kg de magnésium (0,06 %) 81
kg de soufre (0,05 %) et 44 kg de phosphore (0,03 %).
Les feuilles, seules, sont bien plus riches en matières minérales
que l'ensemble de la biomasse (2,11 % d'azote, 1,56 % de calcium, et 1,04 %
de phosphore, notamment). Fort heureusement, elles retombent au sol, chaque
année, ou périodiquement, et lui restituent leurs éléments
minéraux qui, plus ou moins rapidement, redeviennent réabsorbables
par les racines. Du reste, d'autres substances, même en quantité
très minime, sont à considérer ; le chlore, le sodium,
le silicium, le fer, l'aluminium, le manganèse, le cuivre, le zinc, etc...
peuvent se révéler utiles à la croissance des jeunes arbres.
On ne sait pas encore très bien à quoi servent, exactement, toutes
ces substances, mais il est évident, par exemple, que l'azote et le phosphore
sont toujours présents dans les noyaux des cellules végétales
(où ils participent à la construction des acides nucléiques
: ADN et ARN) et qu'ils interviennent, on l'a vu plus haut, dans les réactions
de la photosynthèse. L'azote, avec le magnésium, sert à
l'édification de la molécule de chlorophylle, sans doute grâce
à la présence, transitoire, du fer. Le zinc joue un rôle
dans la synthèse d'un acide aminé, le tryptophane, qui conduit
à l'hormone naturelle ou AIA, facteur de croissance indispensable aux
végétaux; le potassium règle, probablement, la perméabilité
des membranes des cellules, etc...
Il est donc indispensable, quand on veut tenter de relier l'activité
photosynthétique à la croissance, de tenter de se placer dans
des conditions de milieu, aussi exactement contrôlées que possible
; l'idéal serait, évidemment, d'utiliser les possibilités
considérables offertes par les phytotrons. Dans la " phytotronic
newsletter " que publie, périodiquement N. DE BILDERLING au Phytotron
de Gif-sur-Yvette, on peut trouver des indications sur les diverses installations
qui, dans le monde entier, s'occupent des jeunes arbres forestiers : de Stockholm
à Oslo, à Moscou et à Irkoutsk, d'Ottawa à Innsbruck
et à Rotorua en Nouvelle-Zélande, leur nombre se multiplie constamment.
Mais, la plupart du temps, étant donné les grandes dimensions
qu'atteignent, rapidement, certaines espèces forestières, on doit
rester dans le milieu naturel, et il faut s'efforcer de se placer dans des conditions,
de sol en particulier, aussi uniformes que possible.
RÔLE DE LA PHOTOSYNTHÈSE DANS L'INSTALLATION
ET LE PREMIER DÉVELOPPEMENT DES ARBRES FORESTIERS.
L'analyse, même très sommaire, des publications, relativement
récentes, qui ont traité de l'installation et de la croissance
des jeunes arbres forestiers entraînerait l'auteur de cette analyse à
des développements d'une ampleur exagérée. On se bornera
à examiner 3 cas précis :
- les recherches faites, sans aucune préparation du sol, en milieu forestier,
- les recherches faites en utilisant des caissettes de végétation,
en forêt,
- les recherches faites en pépinières, l'éclairement étant
gradué grâce à l'emploi de grandes cages de bois, ou de
fibres de verre, et dont les lattes sont diversement espacées.
Recherches faites, sans aucune préparation du sol, en milieu forestier.
Dans l'exemple qui va être donné, pris dans les forêts résineuses
du Haut Doubs, vers 800/1.000 m d'altitude, on a utilisé pour mesurer
l'éclairement des diverses stations, des cellules photoémissives
(luxmètres usuels, sensibles aux seules radiations visibles) ; les mesures
étaient faites vers le milieu de la journée, soit, par temps couvert,
au point d'observation lui-même, soit, par temps ensoleillé, avec
ciel sans nuages, mais dans ce cas en prenant la moyenne de 65 mesures effectuées
sur un carré de 10 m x 10 m, avec le point d'observation à l'intersection
des diagonales. Les valeurs étaient exprimées par le rapport entre
l'éclairement lumineux mesuré sous bois, et celui mesuré,
à peu près au même moment, sur une surface très largement
dégagée : ce rapport est l'éclairement lumineux relatif
de la station (ELr, ou plus simplement Er).
On partait de la lisière, exposée au sud, et, tous les 10 m,
on mesurait l'Er régnant à environ 1 m de hauteur au-dessus du
sol ; le peuplement étudié était composé de résineux
communs dans cette région (Abies pectinata et Picea excelsa),
âgés d'une centaine d'années, et implantés d'une
façon régulière (521 tiges et 729 m3 par ha).
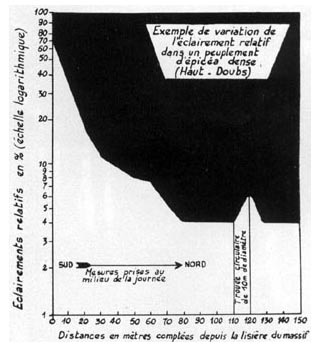
L'Er était voisin de 65 %, à l'alignement des arbres de lisière,
et il s'abaissait régulièrement jusqu'à 4 % environ, vers
70 ou 80 m de cette lisière. Sauf dans de petites trouées, où
l'Er remontait un peu, ou, au contraire, dans des emplacements plus sombres,
où il descendait à 3 et même 2 %, les valeurs relevées
restaient très proches de 4 %.
Quatre ans avant ces observations avait eu lieu une production très
abondante de semences d'épicéas, et les semis s'étaient
installés partout, d'une façon bien uniforme ; comme les bonnes
années de fructification de cette espèce sont assez espacées,
dans cette région, il était donc facile de suivre le développement
des sujets, et l'on a pu établir une différence considérable
de dimensions, entre ceux qui s'étaient développés dans
les 25 à 30 m de la lisière (jusqu'à un Er d'environ 10
à 15 %), et ceux situés plus profondément dans le massif,
ou bien à l'intérieur de sous-étages assez fournis. Comme
les résineux étudiés perdent, très rapidement, leur
faculté phototropique, l'effet de lisière ne produit aucune déformation
de l'axe principal ; c'est donc la hauteur qui a été choisie comme
test de croissance, et l'on a pu observer, approximativement que cette hauteur
variait comme le logarithme de l'éclairement relatif. Au voisinage de
4 %, et au-dessous, la survie n'était plus assurée, et au bout
de quelques années, on a observé, en effet, que la majorité
des sujets avait disparu, faute d'une lumière suffisante.
Ces observations étaient faites sur un sol de rendzine calcaire, profond
et fertile, sans aucun affleurement rocheux ; les précipitations printanières
et estivales sont, dans cette région du Jura régulières
et abondantes, et la régularité du peuple ment supérieur
incitait à penser que la concurrence radiculaire devenait, dès
que l'on s'écartait de la lisière, assez uniforme. On pouvait
donc estimer que les variations de l'éclairement, en agissant sur l'allure
de la photosynthèse, déterminaient d'une façon stricte
le développement des jeunes sujets. En fait, des expériences faites
ultérieurement, en cases de végétation à divers
types de sol, l'éclairement étant mesuré, cette fois, en
continu, ont apporté les mêmes conclusions et montré que,
dans des cas analogues, la méthode utilisée avait une valeur pratique
suffisante. L'évaluation de l'éclairement par la méthode
des photographies hémisphériques (BECKER), conviendrait très
bien à ce type de recherches.
Recherches faites en utilisant des caissettes de végétation installées
en pleine forêt.
Les arbres feuillus, les Fagales en particulier qui ont, en France une grande importance économique, sont, pendant de nombreuses années, très sensibles à l'action de la lumière latérale ; il est donc difficile de se placer, pour les étudier, dans des lisières, comme dans le cas précédent. Le type de recherches à effectuer, dans la moyenne vallée de la Saône, avait un but assez précis : tenter de déterminer, pour Quercus pedunculata et Quercus sessiliflora, le point de compensation pratique, c'est-à-dire l'éclairement minimal au-dessous duquel les matières oxydées par la respiration étaient plus importantes que celles produites par la photosynthèse, ce qui entraînait la disparition rapide des semis naturels. Dans cette région, les glandées abondantes ne sont pas très fréquentes (tous les 6 ou 8 ans, par exemple, mais avec de grandes irrégularités) et il importait d'en tirer parti, au mieux, afin de développer les ensemencements naturels en chênes, dans des forêts traitées souvent en conversion de futaie. Or, on observait assez souvent que des régénérations naturelles, d'apparence bien florissantes, disparaissaient rapidement, même si on les mettait en lumière pendant l'hiver qui suivait leur installation ; on pouvait se demander donc si la nature du sol n'intervenait pas aussi dans ce cas, et il fallait opérer avec certaines précautions. Par ailleurs, comme le peuplement supérieur était très irrégulier, et que l'on devait rechercher divers types de couverts naturels, on pouvait craindre que la concurrence radiculaire des grands arbres en place ne vienne perturber sérieusement, dans certains cas, l'alimentation en eau des jeunes sujets. Il était donc indispensable de recourir à des caissettes de végétation, remplies d'un sol uniforme et soustraites, très efficacement, à la concurrence des racines voisines, pendant les premières années tout au moins.
 |
 |
FIG. 8. - Variations de l'ELr, quand on pénètre dans une futaie résineuse dense du Haut Doubs (521 liges et 729 m3 par hectare), et relations observées entre ces variations et le développement des régénérations naturelles de Picea excelsa, à l'âge de 4 ans.

Lamto. Côte-d'Ivoire. Photo hémisphérique
de forêt riveraine du Bandama.
Photo Bony.
On a donc installé 6 de ces caissettes dans 6 emplacements choisis,
des taillis sous futaie très denses à la très grande trouée,
largement dégagée, et assimilée au plein découvert.
Pour la mesure de l'éclairement, on a installé, près de
chaque caissette, un pyranomètre totalisateur de Bellani, enregistrant
l'ensemble des radiations provenant du soleil et ciel : ces appareils étaient
relevés chaque jour, dans les stations bien dégagées, et
chaque semaine, dans les stations les plus sombres ; les observations ont été
poursuivies pendant 2 années complètes, en continu pendant le
printemps et l'automne, par périodes seulement, en été
et en hiver (quand le couvert reste stable).
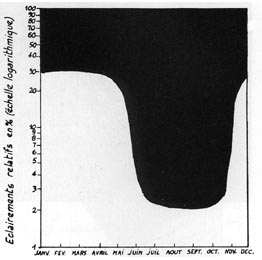
Les résultats furent exprimés par le rapport entre l'éclairement énergétique de la station, et celui reçu dans la grande trouée, très largement dégagée : c'est l'éclairement énergétique relatif ou EEr (désigné plus simplement par Rr). Les résultats obtenus ont été les suivants, pendant la période de végétation active: 3,5 %, 4,3 %, 13,5 %, 57 %, 66 %, et, naturellement 100 %. En même temps, des mesures instantanées répétées d'éclairement étaient faites au voisinage des 2 stations les plus ombragées, et l'on relevait, sur quelques mètres de distance, de légères variations (de 2 % à 5 % de Rr, approximativement). Une caissette, en outre, était abritée par une claie très faiblement ajourée, et recevait au sol environ 1 % de Rr.
 |
 |
|
|
Etat des semis au bout de 2 ans |
Etat des semis au bout de 2 ans
sous un EEr de 100 % en été.
|
|
 |
 |
|
|
État des semis au bout
de 2 ans
sous un EEr de 3,5 % en été. |
Variation de l'EEr, au cours d'une
année complète,
sous un taillis sous futaie très dense. |
FiG. 9. -Expériences pour la détermination du point, de compensation
pratique de deux espèces de chêne, dans un taillis sous futaie
dense de la moyenne vallée de la Saône.
Des glands de Quercus pedunculata et de Quercus sessiliflora furent
mis dans chaque caissette, à l'automne, et recouverts de quelques feuilles
mortes, pour se rapprocher au maximum des conditions existant dans le milieu
naturel.
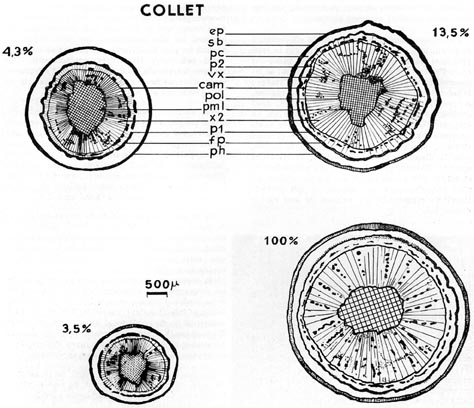
FiG. 10. - Coupes effectuées au niveau du collet,
sur des sujets de Quercus pedunculata, prélevés après
le 2e été, après développement sous des
EEr de 3,5 %, 4,3 %, 13,5 % et 100 %.
ep : épiderme ; sb. : suber ; pc : parenchyme cortical ; p2:
phloème secondaire ; vx: vaisseaux ligneux ; cam : cambium ; pol :
pôle ligneux ; pml : parenchyme médullaire lignifié ; x2
: xylème secondaire ; p1 : phloème primaire ;
fp : fibres péricycliques ; ph : phloème (A. TRONCHET et J. GOGUELY).
Dès le mois d'Avril suivant, on a constaté, dans toutes les caissettes,
une germination très régulière (taux de 80 %) et les jeunes
sujets se sont mainte nus partout en bon état, tout au moins en apparence.
Même sous 1 % de Rr, les petits chênes semblaient en parfaite santé,
mais un essai simultané de germination, dans une obscurité complète,
donnait des tiges très minces et très longues, décolorées,
avec les feuilles réduites à l'état de petites écailles.
Tous ces résultats, bien entendu, s'expliquaient très bien par
les réserves abondantes existant dans les glands, qui rendaient la germination
indépendante de la photosynthèse. Des examens plus poussés
montraient cependant des différences nettes de structure entre les sujets
développés dans les diverses stations. Lors du printemps suivant,
tous les sujets ont développé leurs feuillages, et effectué
leur accroissement en longueur, mais, ceux situés sous un Rr de 3,5 %,
et au-dessous, perdaient progressivement leurs feuilles, dépérissaient
visiblement, et, à l'automne, étaient presque tous morts. Les
hampes, représentant les anciennes tigelles, restaient cependant bien
visibles et persistaient pendant plusieurs années. À peine au-dessus
de 4 %, la survie, assez pénible, semblait cependant assurée,
et les sujets se sont maintenus, pas très florissants, pendant quelques
années.
Des observations analogues ont été effectuées, à
nouveau, lors de glandées abondantes qui se sont produites, par la suite,
dans des forêts du Nord-Est de la France, sur sol travaillé ou
sur sol nu, et les conclusions ci-dessus se sont trouvées confirmées
: le point de compensation pratique, dans ces régions, est voisin de
l'éclairement relatif de 4 % (soit environ 16 calories par cm2
et par jour moyen, en été). Des observations microscopiques très
nombreuses ont été effectuées, sur les sujets des diverses
stations, par l'Institut Botanique de Besançon, sous la direction de
A. TRONCHET, et elles ont apporté diverses précisions sur les
anomalies de structure que présentaient les jeunes chênes les plus
sous-alimentés en lumière, et qui permettaient de comprendre les
raisons de leur rapide disparition.
Recherches faites en utilisant des cages aux parois diversement ajourées.
Il n'est pas toujours facile de se procurer, d'installer, et de relever régulièrement
des appareils enregistreurs, ou totalisateurs de lumière, ou de radiations
solaires ; on peut, évidemment utiliser des méthodes plus rapides,
comme les photographies hémisphériques, on l'a déjà
indiqué. Mais, depuis de nombreuses années, certains chercheurs
se servent de dispositifs (films de plastique, claies, cages, etc...) qui simulent
plus ou moins exactement les couverts naturels. E. BJÖRKMANN (1945) a rapporté
les résultats d'expériences de longue durée, entreprises,
dès l'année 1932, dans le nord de la Suède (au voisinage
du 64° parallèle) sur de jeunes sujets de Pinus sylvestris, provenant
de régénérations naturelles surtout, et placés pendant
de nombreuses années (9 ans, et 5 ans) dans de grandes cages de bois
(réduisant l'éclairement à 50 % et à 25 %), ou sous
des claies (réduisant l'éclairement à 50%). Les résultats,
difficiles à interpréter en raison des rigueurs du climat dans
ces régions assez voisines du cercle polaire, montrèrent, surtout,
qu'il fallait éviter tout changement trop rapide dans les conditions
d'éclairement naturel des sujets.
Plus récentes sont les expériences de K. T. LOGAN, au Canada,
dans la station de recherches forestières de Petawawa, au voisinage du
Lac Ontario, commencées en 1957, avec 22 espèces, résineuses
et feuillues, dans une vaste pépinière ; l'ombragement était
obtenu grâce à de grands abris en lattes et fibres de verre laissant
passer respectivement : 13 %, 25 % et 45 % de la lumière du plein découvert.
Le sol, uniforme, était une argile sablonneuse, et les sujets étaient
arrosés pendant les périodes de sécheresse estivale, surtout
en plein découvert. Des mesures d'humidité du sol, effectuées
par moments, montraient que le sol était plus humide dans les stations
les plus ombragées que dans celles qui étaient les plus découvertes.
Le pH variait extrêmement peu entre les divers emplacements. Des mesures
intermédiaires des sujets étaient effectuées, mais, c'est
au bout de 9 ans que les résultats les plus précis ont été
obtenus.
Par exemple, pour 4 résineux : Picea mariana, Picea glauca, Abies
balsamea et Thuya occidentalis, on a trouvé que la hauteur
totale des sujets augmentait régulièrement jusqu'à 45 %
d'éclairement relatif, puis alors que la croissance augmentait plus faiblement,
pour les 2 premières espèces, dans des éclairements supérieurs,
elle diminuait un peu pour les 2 dernières. Mêmes constatations
en ce qui concernait le diamètre de la racine au collet.
En ce qui concerne le poids sec de la seule tige, et le poids de l'ensemble
de la partie aérienne, on constatait une augmentation régulière
avec l'éclairement relatif, jusqu'à une valeur de 45 %, puis,
alors que cette augmentation se poursuivait moins rapidement pour les 3 premières
espèces, jusqu'au plein découvert, il diminuait pour la 4e
; peut-être les arrosages d'été n'étaient-ils pas
suffisants pour cette espèce d'ombre, caractérisée du reste
par un feuillage très abondant (en poids, à 9 ans, il représentait
88 % du poids total de la partie aérienne). En tout cas, l'éclairement
relatif de 45 % semblait constituer un point d'inflexion dans l'allure de la
croissance de toutes les espèces en expérience.
En conclusion de ces trois types de recherches, on peut conclure que, du point
de vue sylvicole, 2 éléments du microclimat lumineux semblent
très importants à déterminer
a) le point de compensation pratique, en milieu naturel, qui correspond
à l'éclairement relatif minimal au-dessous duquel les jeunes arbres
ne peuvent survivre, leurs pertes par la respiration devenant supérieures
à leurs gains, par la photosynthèse ;
b) le point de saturation pratique, en milieu naturel, qui correspond
à l'éclairement relatif maximal, au-dessus duquel on ne constate
plus de gain appréciable de croissance, la photosynthèse étant
alors freinée par suite de la déficience de certains autres facteurs
du milieu ; il s'agira alors souvent de l'eau disponible dans le sol.
Ces deux éléments, exprimés de préférence
en valeurs relatives, peuvent être déterminés : le premier,
grâce à des luxmètres, pyranomètres, etc., à
des cages, ou par la méthode des photographies hémisphériques
; le second, par les procédés qui viennent d'être indiqués,
ou bien par une méthode théorique adaptée par R. CATINOT
aux régions équatoriales et qui sera souvent suffisante.
RÔLE DE LA PHOTOSYNTHÊSE DANS L'ACCROISSEMENT
DES PEUPLEMENTS FORESTIERS CONSTITUÉS.
La photosynthèse étant le support indispensable de la croissance
des peuplements constitués, il doit exister certaines relations entre
leur surface foliaire, et leur accroissement courant, ou moyen annuel. Mais,
pour les raisons exposées plus haut, ces relations ne sont évidentes
que statistiquement, si l'on considère un certain nombre de types de
peuplements, et ce, pendant un certain nombre d'années. La mesure de
la surface foliaire est donc importante : c'est un problème qui, en lui-même,
ne présente pas de difficultés techniques spéciales, à
condition, évidemment, de s'armer de beaucoup de courage et de patience
!!! Du reste, à côté de cette surface proprement dite, d'autres
paramètres doivent intervenir : si l'on admet que la lumière qui
agit sur la photosynthèse a une direction principale verticale (ce qui
n'est vrai qu'approximativement), il faut considérer l'inclinaison moyenne
de l'axe de la feuille (ou de l'aiguille) par rapport au plan horizontal, et
également, le degré d'ouverture ou de fermeture du limbe, uniquement
cette fois pour les arbres feuillus, de part et d'autre de cet axe.
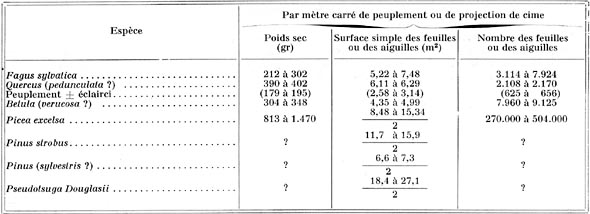
En ce qui concerne la mesure de la surface foliaire des arbres isolés, ou des peuplements, après divers résultats plus ou moins fragmentaires, il semble que ce soit C. MOLLER qui, en 1945, ait publié le travail le plus important, sur des dénombrements opérés dans les forêts danoises, mais surtout du reste, sur Fagus sylvatica et Picea excelsa : les chiffres varient, naturellement, avec chaque espèce, chaque qualité de station, chaque âge et chaque densité de peuplement, et le tableau ci-dessus montre à quels " ordres de grandeurs " on peut s'arrêter.

Poussée d'un jeune Niangon mis en lumière par
des travaux d'amélioration des peuplements naturels en CÔte d'Ivoire.
Photo Lepître.
L'angle d'inclinaison des feuilles sur l'horizontale, et le coefficient d'ouverture
du limbe ont été étudiés spécialement par
GRULOIS (1966), dans la chênaie-hêtraie à charme de Virelles
Blaimont, en Belgique. Voici quelques valeurs obtenues à la suite de
ces observations, dirigées par A. GALOUX.
Pour Quercus pedunculata, l'angle d'inclinaison des feuilles, sur l'horizontale
varie de 29° 7, au sommet de la cime, à 18° 1 dans la partie
basse de celle-ci. Le degré d'ouverture varie de 7,35 % au sommet de
la cime (feuille un peu repliée), à 2,32 % à la base de
celle-ci (feuille nettement plus étalée).
Pour Fagus sylvatica, l'angle d'inclinaison des feuilles, sur l'horizontale,
varie de 33° 2, au sommet de la cime, à 7° dans la partie inférieure
de celle-ci. Le degré d'ouverture du limbe varie de 17,14 % au sommet
de la cime (feuille déjà bien repliée), à 0,37 %
à la base de celle-ci (feuille pratiquement plane).
Photosynthèse, surface foliaire, et accroissement en poids de la biomasse.
On admet, en général, que l'affaiblissement du rayonnement naturel,
quand il pénètre dans le milieu forestier, obéit à
la loi de BEER, modifiée pour être adaptée aux caractéristiques
de ce milieu. Au niveau du sol, en particulier, on a :
EF = EO e-kF ,
relation dans laquelle EF représente l'éclairement
énergétique au niveau du sol EO l'éclairement
énergétique à la surface supérieure du peuplement,
e la base des logarithmes népériens, F la surface
foliaire simple, et k le coefficient d'extinction du milieu. On a surtout
attribué les variations de k à la plus ou moins grande
inclinaison des feuilles sur l'horizontale, et au plus ou moins grand degré
d'ouverture de leurs limbes. Or, il faut observer qu'en appliquant à
une hêtraie, par exemple, les éléments déterminés
ci-dessus, on arrive à une " surface moyenne projetée
" sur le plan horizontal de plus de 5 ha par hectare de peuplement.
Si ces feuilles étaient jointives, et exactement superposées,
on n'enregistrerait presque plus aucun éclairement au niveau du sol (1
feuille de lumière et 1 feuille d'ombre superposées ne laissent
plus passer que 2/1.000° de la lumière incidente). Or, les mesures
faites dans de telles hêtraies donnent des valeurs d'éclairement
relatif de l'ordre de 3,4 ou 5 % par exemple ; en levant les yeux, on perçoit
parfaitement que le couvert est percé de nombreux orifices par lesquels
pénètrent les rayons lumineux et la méthode des photographies
hémisphériques est basée, précisément, sur
cette discontinuité du couvert. Un élément important, sur
lequel on n'insiste que très peu, est la façon dont les feuillages,
quelle que soit leur surface totale, sont groupés : partant de cette
constatation simple, on peut donc imaginer des peuplements théoriques
dans lesquels les feuilles sont placées sur des plans horizontaux successifs,
mais de façon discontinue, permettant à chacun d'entre
eux, et jusqu'au niveau du sol, de recevoir une partie de l'éclairement
qui atteignait le plan immédiatement supérieur.
Dans l'exemple qui va être donné, une hêtraie, on retiendra
les éléments suivants : les feuillages sont répartis sur
6 plans horizontaux, et ils occupent, sur chaque plan, la moitié de la
surface totale disponible.
L'éclairement moyen qui règne, dans l'Europe de l'ouest, à
la surface supérieure des forêts, peut être estimé
à 30.000 lux environ ; le 1er plan reçoit donc un éclairement
moyen de 30.000 lux, sur la moitié de sa surface, le second de 15.000
lux, le 3e de 7.500 lux, etc...
L'activité photosynthétique de chaque plan est déterminée en considérant, conformément à ce que l'on trouve dans les études élémentaires de la photosynthèse, que sous 30.000 lux, un dm2 de feuillage assimile, par heure, environ 12 mg de dioxyde de carbone, sous 15.000 lux 9 mg environ, etc..., en se raccordant progressivement aux valeurs trouvées pour les feuillages d'ombre. Tout le système fonctionne pendant un nombre d'heures estimé, arbitrairement, à 2.100 par an (5 mois de végétation active). Le chiffre total trouvé correspond à l'assimilation dite " nette " (c'est-à-dire, déduction faite de la respiration diurne des feuillages, qui semble, de beaucoup, la plus importante). On arrive ainsi, par hectare, à une assimilation nette de dioxyde de carbone de l'ordre de 35 t par an (soit l'équivalent approximatif de 21 t de polyholosides : cellulose, lignines, amidon, etc...) ; mais la biomasse ne s'accroît pas d'un tonnage aussi important, car, dans les évaluations ci-dessus, on n'a pas tenu compte encore des pertes supplémentaires dues à la respiration nocturne des feuillages, ainsi qu'à celle, permanente, du surplus du corps des arbres du peuplement : les troncs avec leurs branches, et les racines. Des divergences assez grandes existent entre les estimations de ces respirations supplémentaires : SCHULZE & KOCK (1971) pensent qu'elle peut représenter 50 % de l'assimilation nette, mais TRANQUILLINI (1959), dans un travail très poussé effectué sur de jeunes résineux, enfermés entièrement dans des enceintes transparentes, où les variations de la teneur de l'air en dioxyde de carbone étaient minutieusement enregistrées, arrivait à des chiffres nettement inférieurs (39 % de l'assimilation brute, soit à peu près 25 % de l'assimilation nette, respiration diurne des feuillages déduite) ; en retenant cette valeur, on arriverait à un accroissement net de la biomasse, en supposant les conditions d'éclairement et d'alimentation en eau convenablement remplies, de 15 à 16 t de matières sèches, par hectare et par an. On n'est pas très loin des estimations de MÖLLER, obtenues par des voles plus directes : 11 à 16 t, pour les hêtraies, ni de celles de DUVIGNEAUD & AMBROES, pour des chênaies, cette fois, de 12 à 15 t.
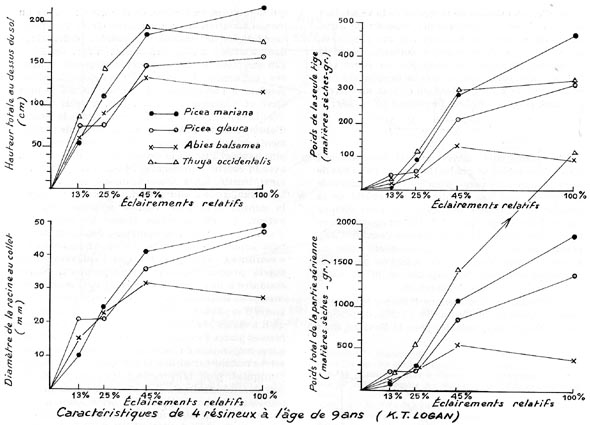
FiG. 11.
Dans cet exemple, très théorique, la surface foliaire projetée
n'était que de 3 ha, mais le calcul montre qu'avec 12 étages de
feuilles, représentant, avec le même espacement, 6 ha de surface
projetée, la production potentielle de la biomasse ne serait majorée
que de 5 % environ ; on peut, bien entendu, procéder de diverses autres
façons, et cet exemple montre seulement que l'on peut, à partir
de considérations relatives à la photosynthèse unitaire
des feuillages, à leur répartition dans l'espace, et à
leur surface totale, et au temps pendant lequel ils fonctionnent effectivement
chaque année, arriver à des conclusions qui ne sont pas trop déraisonnables.
Photosynthèse, absorption de l'énergie lumineuse par les cimes,
et accroissement en bois fort des peuplements forestiers.
On peut reprendre ce problème d'une façon plus détaillée, en faisant intervenir la densité des peuplements qui entraîne, incontestablement, une variation dans l'importance de la masse foliaire, laquelle, on l'a vu, conditionne l'allure de la photosynthèse. On peut d'abord établir que la quantité de radiations absorbées par les cimes est d'autant plus élevée que le peuplement est plus dense, ce qui est évident, mais aussi qu'elle se fait selon certaines lois qui relient ces deux éléments entre eux d'une façon plus précise. À la base des études de l'absorption de l'énergie lumineuse par les cimes de peuplements de densité différente est la mesure, expérimentale, du pourcentage de cette énergie reçu au niveau du sol du sous-bois : toutes les relations obtenues entre ces deux éléments sont matérialisées, graphiquement, par des branches d'hyperboles. La partie située au-dessous de la courbe représente l'énergie transmise, et la partie située au-dessus, l'énergie retenue par l'ensemble des cimes ; on a trouvé que, pour les peuplements feuillus, environ 1/3 de l'énergie absorbée est retenu par les seuls feuillages, les 2/3 restants sont retenus par les branches et le tronc, qu'ils contribuent surtout à réchauffer, au moment de la reprise de la végétation annuelle. Pour les peuplements résineux du Jura, où diverses séries de mesures ont été effectuées, on a pu proposer la relation suivante, entre la proportion d'énergie absorbée par les cimes (Q), l'albédo de la surface supérieure du peuplement (A), le nombre de tiges par hectare (N), et un facteur constant pour ces espèces forestières (K = 20) :
![]()
D'un autre côté, divers auteurs forestiers ont établi des
" tables de production " pour certaines de leurs espèces forestières
principales. Ces tables de production sont des " tables de probabilité
de la production ligneuse " qui donnent, pour chaque espèce,
pour chaque qualité de station, pour chaque âge du peuplement et
pour chaque densité d'implantation des arbres, l'accroissement courant
annuel, ou moyen annuel, sur lequel on peut vraisemblablement compter, exprimé
en m3 de "bois fort ", de 7 cm et plus de diamètre.
En Allemagne, E. ASSMANN (1965) a établi, par exemple, pour Picea excelsa,des " tables de production " très précises, valables pour la Bavière, mais qui sont citées, ou copiées, dans toute une partie du monde forestier. Cet auteur envisage de très nombreux types de stations (33 qualités différentes), dans chacune d'elles des classes d'âge, de 20 à 120 ans (tous les 5 ans), et divers degrés de densité des peuplements (de 0,4 à 1,2 ; la densité 1 correspondant à l'accroissement courant annuel le plus élevé et supérieur d'environ 3 % à celui obtenu avec la densité de 1,2). Or, en comparant la courbe d'absorption des radiations par les cimes de peuplements de densité croissante, ainsi qu'on l'a expliqué plus haut, et jusqu'à la densité maximale observée, avec la courbe représentant la relation : accroissement/densité, selon ASSMAN, on constate une certaine similitude d'allure. Ceci, sauf au voisinage de la densité considérée comme optimale par cet auteur, qui est lui-même étonné de cette sorte d'anomalie puisqu'il se demande, dans l'un de ses plus importants ouvrages, " Waldertragskunde " : " comment est-il possible que l'enlèvement de sujets productifs, dans un peuplement, puisse conduire à une augmentation de son accroissement ? ". Il élabore donc diverses hypothèses pour tenter d'expliquer, physiologiquement, cette relation qu'il a établie, expérimentalement, sur de très nombreuses places d'expérience ; il pense, naturellement, à une majoration de l'activité photosynthétique des arbres restant sur pied, à la suite des coupes légères pratiquées, pour faire revenir les peuplements de leur densité maximale (1,2 environ) à leur densité considérée comme optimale (I). Selon ASSMANN, On pourrait obtenir ainsi une petite augmentation de la teneur en dioxyde de carbone de l'air des sous-bois, une amélioration des conditions de transpiration, maintenant plus longtemps les stomates des aiguilles ouverts, etc... On peut penser aussi que, dans ces peuplements " travaillés ", les aiguilles d'ombre sont, périodiquement (au moment des éclaircies) légèrement suréclairées, et, comme elles ont une activité photosynthétique un peu supérieure à celle des aiguilles de lumière, il pourrait en résulter un accroissement, assez minime, de l'ensemble de la nutrition des arbres demeurant sur pied.
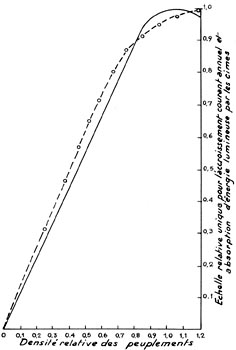
FIG. 12. - La courbe représentant les relations
entre la densité d'un peuplement et son accroissement courant annuel,
et la courbe représentant les relations entre la même densité
et l'énergie lumineuse absorbée par les cimes, ont une certaine
similitude d'allure :
_____ Peuplement régulier de Picea excelsa, âgé
de 50 ans, dans une station de Bavière de très bonne qualité
(classe 0,36-ME selon E. ASSMANN). La densité optimale (1,0),
inférieure à la densité maximale (1,2) est celle qui permet
l'accroissement courant annuel le plus élevé (1,0).
o - - - o Peuplement régulier de Picea excelsa, âgé
de 50 ans, dans une station du Jura d'excellente qualité. La densité
maximale (1,2) correspond à l'absorption maximale d'énergie lumineuse
(1,0).
N. B. - Remarquer la divergence des courbes, au voisinage de la densité optimale d'E. ASSMANN.
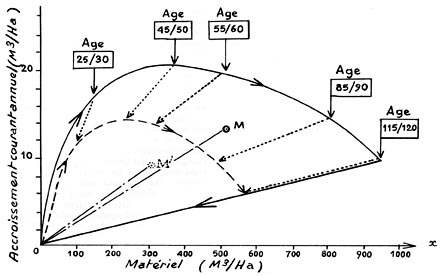
FIG. 13. - Relations entre le matériel sur pied et l'accroissement
courant annuel d'un peuplement régulier de Picea excelsa, de son origine
à son exploitation totale, dans une station de très bonne qualité
(classe : 0,36-ME, selon E. ASSMANN) :
____ Densité optimale.
- - - - Densité relative égale à 60 % de la
densité optimale.
En constituant deux forêts théoriques aménagées
(par exemple : 12 classes d'âges : 1 à 10 ans, 11 à 20 ans,
etc.... réparties sur des surfaces égales), on obtient :
en M, la relation entre le matériel et l'accroissement courant
annuel pour l'ensemble des classes d'âge, à la densité optimale,
en M', la relation entre le matériel et l'accroissement courant
annuel pour l'ensemble des classes d'âge, à une densité
égale à 60 % de la densité optimale.
Les tangentes des angles MOx et M'Ox matérialisent, respectivement, les taux d'accroissement de ces deux types de forêts.
Les tables de production, en général, peuvent donner bien d'autres
renseignements, sur lesquels on n'insistera que très peu :
1) Une forêt aménagée étant constituée par
une série de peuplements d'âges croissant, on peut constituer des
forêts aménagées théoriques, et calculer l'accroissement
courant annuel, ou moyen annuel total : par exemple, avec les tables
d'ASSMANN, on peut montrer que, dans une station très uniforme, une forêt
aménagée d'épicéa dont le matériel moyen
sur pied ne sera égal qu'à 60 % du matériel considéré
comme optimal, aura un accroissement courant annuel égal à 70
% environ de celui obtenu avec le peuplement de densité optimale. Le
facteur " densité ", sur lequel les sylviculteurs peuvent souvent
agir, est donc l'un des éléments importants de la production ligneuse.
2) Si l'on considère, par contre, le volume moyen de l'arbre produit,
dans les deux forêts théoriques décrites ci-dessus, on trouvera
qu'il est plus élevé dans le premier cas (forêt un peu allégée),
que dans le second cas (forêt à densité optimale). Le taux
d'accroissement du matériel sur pied sera également un peu plus
fort dans le premier cas.
3) On constate, très généralement, que l'accroissement
courant de chaque peuplement régulier, quel que soit l'auteur de la table
de production, passe par un maximum à un âge, en général
peu avancé dans les stations de très bonne qualité (40
à 50 ans pour Picea excelsa ; 50 à 60 ans, pour Quercus
sessiliflora) ; quand la qualité de la station se dégrade,
l'âge augmente dans d'assez grandes proportions. On en a déduit,
un peu hâtivement, que les arbres de ces peuplements perdent assez
vite leur activité de croissance, et qu'ensuite, ils végètent
plus ou moins péniblement, jusqu'à ce que les forestiers se décident
à les réaliser. Cette conclusion est inexacte. En effet, si l'on
calcule l'accroissement de l'arbre moyen restant sur pied, dans les divers
types de peuplements étudiés, on constate qu'il ne cesse d'augmenter
avec l'âge, ainsi que ceci avait été avancé par HÜFFEL,
il y a plus d'un demi-siècle. Pour Picea excelsa, par exemple,
les tables d'ASSMANN montrent qu'à l'âge de 20/25 ans, l'arbre
sur pied s'accroît de 0,003 m3 par an, à l'âge
de 60/65 ans, de 0,019 m3 par an, et à 115/ 120 ans, de 0,024
m3 par an (soit 8 fois plus qu'à 20/25 ans). Pour Quercus
sessiliflora, les tables de PARDÉ-DECOURT donnent à 30 ans,
pour un arbre sur pied, un accroissement annuel de 0,003 m3, - à
60 ans, de 0,011 m3, - à 90 ans de 0,017 m3, -
à 120 ans, de 0,028 m3, - à 180 ans de 0,040 m3,
et à 220 ans, près de la limite d'âge retenue par ces tables,
de 0,137 m3. Or, plus l'arbre se développe, plus la surface
de ses feuilles s'accroît, et plus il est isolé, plus l'éclairement
de sa cime est intense ; il est donc très logique que son activité
photosynthétique soit fortement majorée, donc que son accroissement
annuel soit de plus en plus élevé, jusqu'à la limite de
sa longévité naturelle, bien entendu. En tout cas, la diminution
de l'accroissement courant annuel d'un peuplement, que l'on constate généralement,
à partir d'un certain âge, n'est pas due, essentiellement, à
la baisse de l'activité physiologique de chacun des arbres qui le constituent,
mais, d'une façon beaucoup plus directe, à la diminution du
nombre des arbres restant sur pied, diminution, en partie inéluctable,
en partie provoquée par les habitudes culturales des forestiers.
On arrêtera là ces commentaires qui prouvent, d'une façon
surabondante, que la production forestière est liée, très
directement, à l'intensité du phénomène de la photosynthèse,
elle-même conditionnée par un certain nombre de facteurs :
la surface foliaire, et le temps pendant lequel elle fonctionne, sa sensibilité
aux variations des éléments microclimatiques au niveau des cimes,
comme aux changements qui peuvent survenir dans les conditions édaphiques
du milieu prospecté par les racines ; c'est par une recherche, minutieuse
et tenace, de chacun de ces éléments essentiels, que l'on pourra,
en définitive, faire progresser cette physiologie végétale
appliquée qui constitue la base de la sylviculture moderne.
BIBLIOGRAPHIE SOMMAIRE
ASSMANN (E.) (1961). -Waldertragskunde. B. L. V. Verlag, München, Bonn,
Wien, 490 pages.
BASSHAM (J. A.) et CALVIN (M.) (1959). -Le cycle du carbone dans la photosynthèse,
Dunod, 112 pages.
BUVAT (R.) (1954). - Chimiosynthèse et photosynthèse, P. U. F.,
208 pages.
CATINOT (R.) (1965). - Sylviculture tropicale en forêt dense africaine.
Bois et Forêts des Tropiques, n°100 à 104, 71 pages.
COSTES (C.) et divers (ouvrage collectif) (1975). - Photosynthèse et
production végétale, Gauthier Villars, 283 pages.
GIESE (A.) et divers (ouvrage collectif) (1964). - Photophysiology, tome 1,
Academic Press, London, New York, 377 pages.
GRULOY (J.) (1967). - Extinction du rayonnement global, tropismes et paramètres
foliaires. Bulletin de la Société Royale de Botanique de Belgique,
tome 100, pp. 315-334.
JACQUIOT (C.) (1964). - La forêt, Masson et Cie, 160 pages.
LARCHER (W.) (1973). - Ökologie der Pflanzen. Ulmer, Stuttgart, 320 pages.
LOGAN (K. T.) (1969). - Growth of tree seedlings as affected by light intensity.
Canadian Forestry Service, Publication n°1256, 12 pages.
MOYSE (A.) (1952-1953). - La photosynthèse. Année biologique,
tomes 28 (pp. 217-293) et 29 (pp. 165-244).
MÖLLER (C.) (1945). - Laubmenge, Stoffverlust, und Stoffproduction des
Waldes. Kandrup et Wunch's Bogtrykkeri-Kobenhavn, 287 pages + tableaux annexes.
RABINOWITCH (E. 1.) (1945-1951). - Photosynthesis and related process (tomes
1 et II). Intersciences publishers, New York.
ROUSSEL (L.) (1972). - Photologie
forestière, Masson et Cie, 144 pages.
SESTAK (Z.), CATSKY (J.), et JARVIS (P. G.) et divers (ouvrage collectif) (1971).
- Plant photosyntetic production. Manual of methods. W. Junk publishers. The
Hague, 818 pages + annexes.
TRONCHET (A.) et GRANDGIRARD (A.) (1956). - L'analyse histométrique
et son application à l'écologie forestière. Ann. Sci. Univ.
Besançon, 2° série, Botanique 8, pp. 3-30.
UNESCO (1971). - Actes du colloque de Bruxelles de l'année 1969. Voir notamment : La chênaie mélangée calcicole de Virelles-Blaimont, en Haute Belgique et Flux et transferts d'énergie au niveau des écosystèmes forestiers, par P. DUVIGNEAUD, A. GALOUX, et divers.