
Article publié en 1966 dans la Revue Forestière Française, n° 11, p 708 - (télécharger la version .pdf - 398 k)
Le Phototropisme Juvénile des Résineux
par Louis Roussel
Quand on parle de phototropisme en matière forestière, on pense
surtout aux arbres feuillus. Dès leur jeunesse, et jusqu'à un
âge avancé, ces espèces ligneuses conservent la propriété
de s'orienter vers la lumière. C'est surtout depuis les classiques expériences
de WENT (1928) que l'on comprend comment s'exerce cette action : les substances
de croissance, ou auxines, qui stimulent et, sans doute, organisent la construction
des tissus végétaux sont détruites, détournées
ou inactivées par certains rayons lumineux (ceux de couleur bleue notamment).
D'où une croissance plus réduite du côté éclairé,
ce qui provoque la courbure de la tige en cours de développement.
Certains auteurs, J. WIESNER (1902) et A. ENGLER (1924) en particulier, ont
bien étudié les effets divers, souvent confondus dans la pratique,
du géotropisme négatif des tiges (qui les incite à croître
verticalement), et de ce que l'on appelait alors " l'héliotropisme
" (qui tend à les courber vers la lumière). ENGLER a effectué
diverses observations et expériences sur ce sujet : il montre que
l'héliotropisme est tellement puissant qu'il peut courber des tuteurs
placés contre les jeunes arbres feuillus en expérience pour essayer,
dans un éclairement latéral, de maintenir leur croissance verticale.
Il a vérifié également que le changement d'orientation
provoqué par l'héliotropisme pouvait répercuter ses effets
jusqu'à des niveaux inférieurs, qui avaient été
atteints plusieurs années avant le changement de direction : ceci
probablement par suite d'un effet de compression et d'extension des tissus ligneux
déjà formés. Le même auteur note que l'héliotropisme
des sapins et des épicéas d'un certain âge semble inexistant;
les quelques observations de ce genre faites sur des pins et des mélèzes
lui paraissent également très douteuses. Par contre, il signale
certains faits, assez rares, d'héliotropisme observés sur les
pousses des arbres résineux jeunes, Quant au géotropisme, il est
évident, dans tous les cas.
Les forestiers habitués aux résineux du nord-est de la France,
au sapin et à l'épicéa notamment, sont absolument d'accord
avec cette prise de position. Ils ne constatent pas d'orientation sensible,
et déterminée par la lumière latérale, sur des sujets
se développant dans leurs stations naturelles - mais ils ont le souvenir
de hêtres, d'érables ou de frênes, qui " filent "
dans les petites trouées, pour gagner, au prix de quelques contorsions,
le niveau supérieur des peuplements. Et ces comportements divers, dans
les " sociétés d'arbres " que sont les forêts,
évoquent souvent pour eux l'attitude des individus dans les sociétés
humaines.
Il est donc, en tout état de cause, intéressant de se poser quelques questions au sujet du phototropisme des résineux : ces espèces ligneuses sont-elles, en toutes circonstances, insensibles à l'action de la lumière latérale ? Quelles pourraient être les raisons physiologiques de leur comportement et quelles conséquences culturales pourrait-on en tirer?
**
Grâce à divers envois de graines effectués par M. BOUVAREL,
Directeur de Recherches au C.N.R.F. de Nancy, en 1962, 1963 et 1966, il nous
est possible de répondre très partiellement à ces interrogations.
Les premiers envois étaient surtout destinés à tenter de
déterminer les exigences en lumière de 12 espèces résineuses
utilisées dans les reboisements français (4 sapins - 4 épicéas
- 2 pins - 1 mélèze et 1 douglas). Mais les graines restant disponibles
ont été utilisées à des essais de germination, en
cases de végétation à seul éclairement latéral,
en 1963 et en 1966. Voici, sommairement décrit, le dispositif adopté :
En 1963, une longue caisse couverte, mais ouverte partiellement d'un seul côté,
remplie d'un sol naturel non fumé (du type pseudogley), a reçu
12 lots de 100 graines chacun. Les espèces utilisées étaient
les suivantes : Abies pectinata (races du jura et de l'Aude) - Abies
grandis - Abies nordmanniana - Picea excelsa (races du jura et des Alpes
du Sud) - Picea omorica - Picea sitchensis - Pinus laricio corsicana - Pinus
strobus - Larix leptolepis - Pseudotsuga douglasii.
Dans cette caisse, qui recevait par un dispositif de chicanes très sommaire
les précipitations naturelles, la répartition, complexe, de la
lumière était difficile à déterminer de façon
précise. Cependant, grâce à des mesures effectuées
par divers types de temps, avec une cellule, il est possible de dire que les
sujets se développant dans la partie la mieux éclairée
recevaient, en lumière horizontale, environ 20 % d'éclairement
relatif vers l'extérieur, et environ 1 % d'éclairement relatif
vers l'intérieur. Ceux se développant dans la partie la moins
bien éclairée recevaient environ 5 % d'éclairement
relatif vers l'extérieur, et environ 0,05 % d'éclairement relatif
vers l'intérieur. De toutes façons, il existait une différence
importante entre l'éclairement régnant sur les faces internes
et externes des jeunes tiges.
Dans le cours de l'été 1963, on a constaté que toutes les espèces manifestaient un phototropisme net ; les axes hypocotylés s'orientaient vers la lumière, ainsi, du reste, que les cotylédons. On ne pouvait invoquer ici une action unilatérale sur le bourgeon terminal, ou sur l'apex, puisque tous ces résineux manifestaient une germination épigée. Cette orientation montrait bien également que les sujets possédaient une auxine sensible à la lumière, d'un genre analogue à celle, bien connue, du coléoptile d'avoine. (En fait, CLARK et BONGA, dès 1962, avaient détecté une telle auxine sur un sapin américain en utilisant la chromatographie et le test Avena). Par ailleurs, se trouvaient confirmées les observations de GALSTON, BRIGGS, et collaborateurs (1949-1964), au sujet de l'action orientante de la lumière sur l'ensemble de l'axe en cours de développement. Toutefois, on doit remarquer que ces auteurs avaient surtout opéré sur des coléoptiles de graminées.
FIG. 1. Phototropisme juvénile de Pinus laricio austriaca (Champagne).

FIG. 2. Phototropisme juvénile de Pinus sylvestris (Champagne).
Dans le cours de l'année 1966, des essais de germination et de croissance en case de végétation à humidification constante, remplie de terre de bruyère et soumise à un seul éclairement latéral, ont été recommencés sur les espèces suivantes : Abies pectinata, race de l'Aude - Picea excelsa, race du Jura - Pinus strobus - ainsi que sur deux nouvelles espèces : Pinus laricio austriaca et Pinus sylvestris. Là aussi, le phototropisme est très net ; mais il revêt un caractère différent suivant les espèces : tiges rectilignes et obliques pour celles du genre Pinus, fortement courbées pour celles du genre Picea, à forme approximativement en S, pour celles du genre Abies. Les photographies n° 1, 2 et 3 sont démonstratives à ce sujet .

FIG. 3. phototropisme juvénile de Picea excelsa (Jura).
Il est à noter également que, pendant leur période phototropique, les jeunes résineux semblent, très logiquement, manifester une réaction d'allongement quand on les abrite circulairement (Photographie n° 4).

FIG. 4. Pendant sa " période phototropique ", Picea excelsa s'allonge en abri circulaire latéral (à droite). Comparer au témoin, non abrité (à gauche).
**
Qu'allait-il advenir de ces petits arbres ? allait-on, artificiellement, obtenir
des sujets flexueux, ou tortillards ? leur évolution était attendue
avec une certaine curiosité. Or, dès l'année 1965 (pour
les semis de l'année 1963), on a remarqué que les sapins développaient,
du côté ombragé, un bourgeon latéral qui donnait
naissance à une pousse verticale, devenant axe principal, et non
phototropique. Cette évolution est observable, parfois, dans la
nature, sur des régénérations de sapin, en lisière,
et sous des couverts denses et bas. Les autres espèces (sauf le mélèze,
disparu presque totalement sous cette lumière insuffisante), semblaient
se redresser simplement ; cependant, on remarquait, en début
de saison, une très légère tendance persistante au phototropisme,
avec, parfois, développement d'un rameau latéral du côté
éclairé. En 1966, le redressement s'est absolument maintenu.
C'est donc, très précocement (dès la 3 ou dès la
4e année) que les résineux en expérience perdraient leur
faculté de s'orienter vers la lumière, ainsi qu'on le constate,
dans la pratique, pour des sujets âgés de quelques années.
En tous cas, la photographie n° 5, prise en fin d'année 1965 (à
la fin de la 3e année de croissance) semble bien démonstrative
à ce sujet.
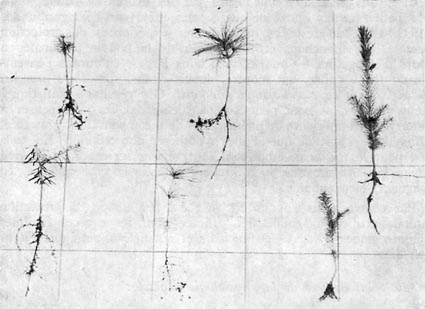
FiG. 5. Sujet de 3 ans d'Abies pectinata (à
gauche) - de Pinus strobus (au centre) et de Picea excelsa (à
droite).
Rangée supérieure : plein découvert.
Rangée inférieure : case de végétation à
seul éclairement latéral, la lumière vient de droite.
Comparer, pour l'épicéa, avec la figure n° 3.
Une exception est cependant à faire pour le pin sylvestre, qui, dans des conditions de souffrance, semble-t-il, et jusqu'à un âge avancé, peut recouvrer cette tendance phototropique, ou devient flexueux, d'une façon assez indéterminée (1).
(1) Toutes ces observations sont, évidemment, effectuées en conditions
d'éclairement naturel (quelques milliers de lux en lumière horizontale)
; on peut les retrouver, dans certains cas, en forêt : jeunes régénérations
de lisière, sous des strates végétales denses, notamment.
Quand on emploie un éclairement artificiel, nettement plus faible, on
arrive à différencier, chez Pinus sylvestris, des groupes
de sujets insensibles à la lumière latérale, et d'autres
qui manifestent un phototropisme plus ou moins marqué. La mesure de l'angle
d'inclinaison est effectuée, comme pour le coléoptile d'avoine,
au bout de quelques jours. La proportion de ces divers groupes de sujets dans
des lots de graines suffirait à caractériser leur origine, et
même à prévoir le comportement ultérieur des jeunes
arbres (inclinaison du tronc, flexuosité, maintien de l'aptitude phototropique,
etc...) (SCHMIDT - KARSCHON).
**
Bien que portant sur un nombre réduit de sujets, ces observations présentent
un certain intérêt :
1° du point de vue de la sylviculture :
Si les résineux, au bout de quelques années, ne manifestent plus
de réaction à la lumière latérale, on peut penser
qu'un abri circulaire (du genre de celui fourni par les arbres de lisière,
dans une petite trouée) sera, lui aussi, sans effets sur l'élongation
des sujets. C'est du reste ce que diverses expériences de protection
latérale (par manchons étroits, autour de la pousse terminale
en cours d'élongation - ou par manchons larges, entourant l'ensemble
du jeune sujet), ont montré, depuis 3 ans, sur Abies grandis, Picea
excelsa et Pseudotsuga douglasii. Ces résultats, statistiquement
négatifs, ne sont peut-être pas définitifs. On peut penser
à l'intervention ultérieure de phénomènes de "
compensatory growth ". Mais il ne s'agit là que d'une hypothèse,
non vérifiée. En tous cas, pour les résineux étudiés,
il faut réviser sérieusement le vieil adage qui affirme que "
la lumière ralentit la croissance ", en longueur, bien entendu.
Par contre, nous l'avons dit (R.F.F., fév. 1965), la protection latérale
artificielle semble très efficace pour les espèces feuillues phototropiques
(Quercus pedunculata et sessiliflora, ainsi que Fagus
sylvatica).
2° du point de vue de la physiologie végétale :
Que de très jeunes résineux soient phototropiques, de la même
façon que des feuillus, jeunes ou adultes, rentre dans le cadre général
des conceptions physiologiques sur le rôle des auxines, leur sensibilité
à la lumière, etc.
Mais que les mêmes résineux, au bout de quelques années,
deviennent insensibles à l'action de la lumière latérale
semble bien moins admissible. D'où vient la perte de cette faculté
? Leur auxine n'est pas modifiée, CLARK l'a montré sur des sujets
plus âgés, et cependant, elle ne parait plus réagir à
la lumière.
Une suggestion, que nous avons déjà formulée, serait que
les formations de protection : l'épiderme avec sa cuticule, et le
suber, même an moment de la croissance terminale et surtout intercalaire,
pourraient par leur opacité, variant avec l'espèce, l'âge
et la station, diminuer tellement la quantité de lumière qui atteint
les couches de cellules en voie de multiplication active et d'allongement que
cette lumière n'aurait plus aucune action freinatrice. Nous avions déjà
émis cette hypothèse, à propos du comportement de deux
chênes (Quercus pedunculata et Quercus sessiliflora) dont
les formations de protection, dans des conditions de station identiques, sont
inégalement transparentes ; la plus opaque correspondant à l'espèce
la plus longue en plein découvert: Quercus pedunculata.
Grâce au microdensitomètre du CNRF de Nancy, très aimablement mis à notre disposition par MM. VENET et POLGE, nous avons pu faire faire quelques mesures de la " densité optique " (exprimée par le logarithme de l'inverse du facteur de transmission optique) des formations de protection des résineux en observation. Pour Abies pectinata, par exemple, l'ancien axe unique semblant, en éclairement latéral, continuer à s'orienter vers la lumière, a des écorces d'une densité optique faible, justifiant ce type de réaction (figure 6 - EC 1 du profil densitométrique). Par contre, l'axe latéral devenant, au cours de la 3e année, non phototropique, a des formations de protection nettement plus opaques (figure 6 - EC 4 du profil, du côté éclairé et EC 3 du côté non éclairé). Ce qui justifierait sa, verticalité, uniquement déterminée par le géotropisme négatif, manifeste dans tous les cas.
En plein découvert, ces formations de protection sont encore bien plus
opaques (figure 6 - EC 7 et EC 8 du profil) ; elles correspondent à l'état
des jeunes sapins développés en pépinière, et installés
ensuite sous un éclairement non équilibré (plantations
de lisières, par exemple) ; on n'observe alors plus aucun phototropisme.
Est-il besoin de préciser que ces idées n'ont encore reçu
que quelques confirmations expérimentales, et qu'il ne convient absolument
pas de les considérer comme scientifiquement établies. Du reste,
même si elles devaient être admises, se poserait le problème
des processus mystérieux qui déterminent la densité optique
différente des formations de protection.
3° du point de vue de la paléobotanique, et, plus spécialement,
de la phylogénèse :
Il est intéressant de remarquer que le phototropisme juvénile et transitoire, de nombreux résineux, se manifeste chez des espèces anciennes très antérieures (souvent de plusieurs dizaines de millions d'années) aux feuillus chez lesquels il se maintient jusqu'à un âge avancé. Or, par ses conséquences (orientation vers la lumière, élongation en cas d'abri naturel bas), cette faculté est profitable à la plante autotrophe, puisqu'elle lui permet de mieux faire bénéficier ses appareils foliacés de la lumière indispensable à sa photosynthèse.

FIG. 6. La densité optique des formations de protection
de segments homologues d'Abies pectinata semble en relation avec leur
phototropisme. Le niveau 3 D indique un facteur de transmission optique de 1/1000e.
(Profils densitométriques de H. POLGE.)
Selon les idées classiques concernant les mécanismes de l'évolution,
ce caractère utile aurait dû apparaître progressivement,
par sélection " à partir du fortuit ". Et l'on devrait
constater que les espèces ligneuses, au fur et à mesure de leur
apparition, présentent un phototropisme persistant de plus en plus marqué.
Or, nous nous trouvons, peut-être, devant l'un de ces cas particuliers
où, selon la définition qu'en donne le biologiste J. ROSTAND,
" ce sont les caractères de l'embryon, ou du jeune - donc transitoires
chez l'ancêtre - qui persisteront chez le descendant jusque dans l'âge
adulte, pour s'intégrer au type normal de l'espèce ".
Cette notion a, déjà, été introduite par H. GAUSSEN
(1937-1945), à propos, précisément, de ses observations
sur l'évolution de divers conifères, et, dans la théorie
très originale (bien que peu connue dans notre pays) qu'il a élaborée,
cet auteur s'exprime ainsi :
" dans une espèce (ou dans un phylum, plus généralement),
un caractère susceptible d'évolution (le phototropisme dans le
cas actuel) apparaît plus développé chez le jeune que chez
l'adulte. Le jeune indique donc le sens de la future évolution "
(2).
Sommes-nous en face de l'un de ces phénomènes, philosophiquement
très intéressants de " coenogénèse prophétique
" ? Doit-on considérer les coniférales comme les lointains
ancêtres des fagales ? Il est trop tôt pour l'affirmer ; mais
cet aspect, très curieux, du phototropisme juvénile des résineux
ne pouvait manquer d'être souligné ici.
(2) Les membres de phrases entre parenthèses ne sont pas de H. GATSEN.