Article publié en 1973 dans la Revue Bois et Forêts des Tropiques, n° 149, p53 - (télécharger la version .pdf - 230 k)
SYLVICULTURE ET PHYSIOLOGIE VÉGÉTALE
Un nouveau venu en physiologie végétale
: le phytochrome
par Louis ROUSSEL.
La croissance des végétaux en général, et des arbres en particulier, s'effectue grâce à la double intervention de matières dites " plastiques " (glucides, lipides et protides), existant dans les semences, lors de la germination - ou bien dans les tissus formés au cours des saisons de végétation antérieures, lors de la reprise de la croissance annuelle - ou encore élaborées lors des multiples réactions de la photosynthèse pendant la pleine période d'activité végétative, et de matières dites " oligodynamiques " de nature variée, qui ont les mêmes origines que ci-dessus, et qui interviennent à très faible dose, comme agents stimulants de la croissance et organisateurs de la matière vivante. Cette distinction très importante est faite par R. DAVID dans son petit ouvrage sur les hormones végétales, lequel, sous réserve de quelques ajustements dus à l'évolution constante, et de plus en plus rapide de nos connaissances, conserve cependant une incontestable actualité (1).
1. - DÉCOUVERTE DE L'EFFET PHOTOPÉRIODIQUE
Les matières oligodynamiques dont il vient d'être fait mention
sont, d'abord, les auxines, et plus particulièrement l'hétéro-auxine
(acide ß indole acétique), agent actif de l'élongation et
de la multiplication cellulaires, puis les gibberellines et les kinines, des
vitamines diverses également, qui toutes interviennent, d'une façon
plus ou moins marquée, pour régler l'allure de la croissance des
végétaux.
Cependant, depuis un certain nombre de décennies, diverses observations
incitaient à penser qu'il existait une autre substance, d'une nature
particulière, qui intervenait pour contrôler le déroulement
d'un certain nombre de phénomènes végétatifs essentiels
: la germination, la croissance et la floraison notamment, ceci en relation
probable avec la durée, variable avec la latitude et le jour de l'année,
du jour et de la nuit.
On peut, évidemment, épiloguer sur les noms des précurseurs,
ou des premiers chercheurs qui s'attachèrent à cerner les contours
du phénomène. Ils sont nombreux, et l'on y relève, dès
l'année 1911, le nom d'un botaniste français, J. TOURNOIS. Mais
on peut admettre que ce sont les expériences de W. W. GARNER et H. A.
ALLARD, publiées en 1920 aux Etats-Unis, qui firent faire à cette
question les progrès les plus décisifs (2).
Ces chercheurs avaient remarqué qu'une variété de tabac
américain (" Maryland Mammoth "), était incapable de fleurir,
en milieu naturel, sous la latitude de Washington. Ils pensèrent que
la durée du jour et de la nuit était à l'origine de ce
comportement spécial, et en organisant une série d'expériences
en serres (jours allongés grâce à un éclairement
complémentaire, ou réduits par l'adjonction d'écrans opaques),
ils démontrèrent l'exactitude de ces vues. Telle semble être
l'origine de la découverte de ce phénomène, que des auteurs
américains continuent à appeler " photopériodicité
", et qui est dénommé chez nous " photopériodisme
".
En multipliant leurs essais, W. W. GARNER et H. A. ALLARD établirent une première classification des plantes, suivant qu'elles fleurissent en jour long, en jour court, ou bien qu'elles sont indifférentes à la longueur du jour (ou de la nuit). Mais, il apparut très vite que ces vues, un peu simplifiées, ne rendaient pas compte de toutes les observations et de nombreux chercheurs s'attachèrent, par la suite, à en déterminer les complexes modalités.
2. - DÉVELOPPEMENT DES RECHERCHES SUR LE CONTRÔLE PHOTOPÉRIODIQUE
DE LA FLORAISON
En France, P. CHOUARD (3), accomplissant un très important travail d'analyse
du contrôle, par la photopériode, de la floraison (1946-1949),
proposa d'admettre la classification suivante, fréquemment retenue chez
les physiologistes de langue française.
- Les plantes de jours longs sont des " héméropériodiques
" (le blé, le seigle, la bruyère, par exemple). Les deux
premières sont des " h. " préférantes, la
troisième est une " h. " absolue, qui ne fleurit absolument
pas en jours courts. La durée du jour long étant nettement supérieure
à 12 heures.
- Les plantes de jours courts, ou plutôt de nuits longues, sont des " nyctopériodiques
". Là encore, on trouve des " n. " préférantes
(comme le soya, variété " Biloxi "), et des " n.
" absolues (comme le tabac " Maryland Mammoth ", objet des premières
expériences de W. W. GARNER et H. A. ALLARD), qu'il est impossible de
faire fleurir en jours longs. Selon P. CHOUARD, la majorité des espèces
tropicales appartiennent à cette seconde catégorie. Le jour court
est, ici, voisin de 12 heures.
- Les plantes indifférentes sont des " aphotopériodiques
" (comme la pomme de terre, par exemple), dont la floraison est indifférente
à la longueur du jour et de la nuit. P. CHOUARD a aussi proposé
de distinguer des plantes " amphiphotopériodiques ", comme
le topinambour, qui demandent, pour fleurir, un ensemble de conditions déterminées
(jours longs, suivis de jours courts, par exemple).
La question du régime photopériodique convenant à la floraison de divers végétaux est devenue très complexe, d'autant plus que l'on a reconnu récemment que certains autres facteurs (comme le froid nocturne, par exemple), pouvaient modifier leurs exigences habituelles : si les nuits sont froides, quelques plantes de jours longs peuvent fleurir en jours courts - et il devient souvent impossible, même en allongeant considérablement la période d'obscurité, de faire fleurir certaines plantes de jours courts (les plantes tropicales, en particulier, selon P. CHOUARD). C. MATHON et M. STROUN, entre autres chercheurs des pays de langue française, se sont employés ces dernières années à approfondir ces notions, qui apparaissent de plus en plus complexes (4).
3. - INFLUENCE DE LA PHOTOPÉRIODE SUR LA GERMINATION ET SUR LA CROISSANCE
Parallèlement à ce genre de recherches concernant la floraison,
se développaient des observations relatives aux conditions de lumière
propres à favoriser (ou à freiner) la germination, ainsi qu'à
celles qui provoquaient l'étiolement.
En ce qui concerne la germination, en ne citant que pour mémoire les
observations de R. CASPARY et de W. KINSEL, il faut arriver à l'année
1935 pour trouver, avec L. H. FLINT et Me E. D. ALISTER la description d'expériences
précises et systématiques, démontrant que la lumière
rouge clair favorisait la germination de certaines graines (laitue " Grand
Rapids "), et que la lumière rouge sombre, lui succédant
immédiatement, inhibait cette germination (5).
On pouvait aussi, du reste, faire certains rapprochements avec les observations
bien plus anciennes de E. VOGT (1915) portant sur un phénomène
très différent : l'étiolement. Ici, le rouge clair supprimait
l'étiolement, alors que l'obscurité le favorisait. Les recherches
concernant l'influence du régime photopériodique, et de la qualité
de la lumière sur la croissance ont été développées,
avec des résultats parfois discordants, dans de nombreux laboratoires.
Assez récemment, J. P. NITSCH, d'abord à l'Université de
Cornell, aux Etats-Unis (1957) puis au Phytotron de Gif-sur-Yvette, a étudié
l'influence de la photopériode sur de nombreuses espèces ligneuses.
Ses observations ont porté sur une quarantaine d'arbustes, ou d'arbres
résineux et feuillus, ornementaux ou forestiers, et certains des résultats
obtenus peuvent intéresser les sylviculteurs. En général,
les sujets se développaient sous des régimes photopériodiques,
partiellement artificiels, ainsi fixés : durée de l'éclairement
de 10 h, 14 h, 18 h et 24 h et durée d'éclairement de 10 h, avec
interruption de la période d'obscurité par 1/2 h d'exposition
à une lumière de composition variée (la plus efficace étant
souvent la lumière fluorescente, rouge clair ou blanche, à peu
près totalement privée d'infrarouges) (6).
Dans certains cas, on trouvait le même développement sous presque
tous les régimes d'éclairement (c'était le cas pour un
if et pour un buis, par exemple). La croissance était vraisemblablement
réglée alors par des rythmes internes, ou endogènes, propres
à chaque végétal.
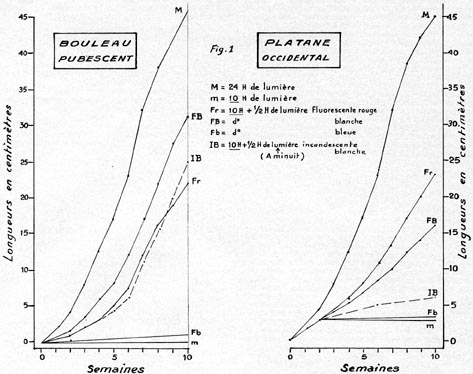
FIG 1.
Dans d'autres cas, par contre, les variations de la croissance étaient
importantes. C'est ainsi que le hêtre commun développait une pousse
de 1 cm de longueur sous un éclairement de 10 h, de 5 cm sous un éclairement
de 18 h, et de 15 cm sous un éclairement de 24 h (en partie artificiel,
il convient de ne pas l'oublier). L'éclairement nocturne complémentaire
de 1/2 h n'avait que très peu d'effet. Ces résultats provenaient
d'expériences poursuivies pendant 3 mois, sur des sujets âgés
d'une année au début de l'expérience. Le pin Weymouth,
âgé de 5 années au début de l'expérience,
réagissait à peu près de la même façon que
le hêtre commun.
L'épicéa commun, âgé de 2 ans, après avoir
fait sa pousse annuelle habituelle, et mis en expérience à ce
moment, reprenait une nouvelle croissance en longueur sous des jours de 18 h
et de 24 h seulement, et aussi sous des jours de 10 h, avec 1/2 h d'éclairement
nocturne.
Le platane occidental, observé dès sa germination, était
sensible à la durée de l'éclairement, et également,
en jours de 10 h, à la 1/2 h d'éclairement nocturne (en lumière
rouge clair surtout). Dans ce cas, le gain de longueur observé, par rapport
au témoin, atteignait 20 cm. Il en était de même pour le
bouleau pubescent, mais, ici, la lumière la plus efficace était
la lumière fluorescente blanche (Fig. 1).
Le pseudotsuga, observé également dès sa germination,
développait une pousse de 2 cm sous des jours de 10 h, de 8 cm sous des
jours de 14 h, de 20 cm sous des jours de 18 h, et de près de 30 cm sous
des jours de 24 h. En jours de 10 h, l'éclairement nocturne complémentaire
de 1 /2 h provoquait un gain de croissance net (10 cm), par rapport aux témoins.
Il apparaissait ainsi, peu à peu, dans l'étude de ces multiples réactions, que la durée du jour et de la nuit, et aussi que l'apport, pendant la nuit, de 1/2 h de certaines radiations, exerçaient une réelle influence sur le développement des végétaux ligneux. Il est à remarquer que les phénomènes de la nutrition carbonée (photosynthèse) ne pouvaient êtres invoqués, car, dans tous les cas, la durée du jour " naturel " était de 8 h 1/2, les compléments, pour arriver aux durées indiquées (10 h, 14 h, 18 h, 24 h et 1/2 h, pendant la période d'obscurité) étaient apportés par des lampes trop faibles pour agir, tant soit peu, sur cette nutrition carbonée nette.
4. - THÉORIES EXPLICATIVES
Entre temps, du reste, avaient commencé, au Laboratoire d'Agriculture
de Beltsville, aux Etats-Unis, les travaux d'une équipe devenue, par
la suite, célèbre : celle constituée par H. A. BORTHWICK,
A. HENDRICKS, V. K. TOOLE et E. H. TOOLE (7). Reprenant, dès l'année
1951, -d'une façon systématique, et avec des moyens importants,
l'ensemble des expériences précédemment indiquées,
ils élaborèrent, peu à peu, la théorie du phytochrome,
qui se présente actuellement de la façon suivante :
Une substance photosensible instable, le phytochrome, existerait en quantité
très faible, chez de nombreux végétaux. Cette substance
a déjà été isolée en 1959 par W. L. BUTLER,
chez de jeunes sujets de maïs. C'est une chromoprotéine (association
d'une protéine et d'un pigment photosensible) de couleur vert-bleu. En
étudiant son spectre d'absorption de la lumière, on a pu la rapprocher
d'un corps chimiquement connu : l'allophycocianine. Ce phytochrome peut revêtir
deux formes : le P. 660, ainsi dénommé parce que sa bande d'absorption
lumineuse principale se situe dans le rouge clair (longueur d'onde de 660 millimicrons),
et le P. 730 (forme active) dont la bande d'absorption principale se situe un
peu au-delà de l'extrême rouge visible (longueur d'onde de 730
millimicrons).
L'interconversion de ces deux phytochromes s'opère suivant le schéma ci-dessous :
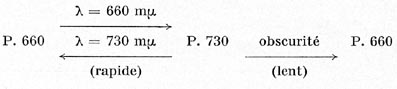
Les réactions rapides, photochimiques, sont en général
peu influencées par la température, tant qu'elle reste dans des
limites normales. Par contre, la réaction lente, qui s'opère à
l'obscurité, dépend assez largement de la température,
comme bien des réactions chimiques ordinaires. Cette remarque permet
de comprendre, tout au moins assez approximativement, le fait déjà
signalé plus haut que les nuits froides peuvent modifier sensiblement
les exigences habituelles de certaines plantes en matière de photopériode.
Selon P. ROLLIN, qui a effectué en 1970 une mise au point très
détaillée sur les multiples publications relatives à ce
phénomène, ainsi que sur ses propres expériences (8), on
peut admettre que, comme les spectres d'absorption de la lumière des
P. 660 et P. 730 se recouvrent partiellement, il existe, sous chaque régime
photopériodique, une proportion déterminée de chacune de
ces deux substances, qui correspond à un état dit " photostationnaire
". Par exemple, en été, sous nos climats, le rapport du P.
730 au P. total (P. 730 + P. 660) serait voisin de 60 %.
Cet état, dans lequel P. 730 serait prédominant, peut provoquer
:
- la germination des graines photosensibles (synthèse d'enzymes diverses,
modification de la perméabilité cellulaire) ;
- le freinage de l'élongation d'un certain nombre de végétaux
(photomorphoses dites négatives), en favorisant peut-être la photo-oxydation
d'un facteur de croissance : l'hétéro-auxine, citée
plus haut;
- l'inhibition de substances encore mal connues (appelées : stimulus
floral, florigènes) qui contrôlent la floraison de nombreuses plantes.
De multiples autres effets peuvent être observés : croissance des feuilles, des racines, mouvements de certains organes, synthèse de pigments divers, etc.... toujours dus à l'intervention du P. 730.
**
En ce qui concerne l'influence de la photopériode sur la floraison,
chez les plantes de jours longs, de jours courts, et indifférentes, A.
HENDRICKS (1964) a proposé d'admettre que les premières produisaient
" trop " de substances florigènes, dépassant ainsi
souvent le seuil supérieur d'inhibition, que les secondes n'en produisaient
normalement " pas assez ", restant ainsi souvent au-dessous du
seuil inférieur d'action, et que les troisièmes maintiendraient
ces substances florigènes dans une, région moyenne d'activité.
On sait, en effet, que bien des substances " oligodynamiques ",
dont il a été parlé plus haut, ne manifestent leur maximum
d'activité que pour un degré de concentration, parfois assez strictement
fixé. Le schéma reproduit ci-dessous, et qui n'est du reste qu'indicatif,
permet de rassembler et de " visualiser " ces diverses notions,
tout en faisant intervenir la température nocturne, dont le mode général
d'action a été décrit ci-dessus. On peut relever, sur ce
schéma (Fig. 2) :
- Que les plantes de jours longs ne fleurissent pas en jours courts, sauf, assez souvent, si l'on apporte un éclairement nocturne complémentaire, ou bien, mais plus rarement, si les nuits sont froides.
- Que les plantes de jours courts ne fleurissent pas en jours longs. En jours
courts, elles ne fleurissent pas non plus, assez fréquemment, si l'on
coupe la période nocturne par un éclairement supplémentaire,
ou bien si les nuits sont froides.
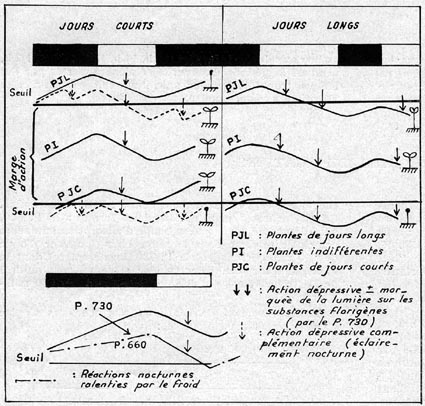
FIG 2.
5. - RÉPERCUSSIONS SYLVICOLES
Il est bien évident que la découverte du phytochrome, parce qu'
elle introduit une série de nouveaux processus en physiologie végétale,
ne peut manquer d'intéresser les sylviculteurs.
La compréhension du rôle joué par ce pigment, dans les
phénomènes de photopériodisme n'est cependant pas, pour
des raisons faciles à comprendre, susceptible de grandes applications
pratiques " actives " en forêt : dans les cultures florales,
il est relativement facile de régler, artificiellement, dans des serres
spécialement aménagées, la durée de la lumière
et celle de l'obscurité. On peut ainsi faire fleurir des chrysanthèmes
en été, par exemple. Dans des pépinières, on peut
envisager, à la rigueur un éclairement nocturne complémentaire.
Mais dans un massif forestier, même de petites dimensions, toute intervention
" active ", et économiquement rentable, parait actuellement
exclue.
Par contre, les forestiers peuvent prendre en considération ce phénomène
pour mieux choisir les espèces nouvelles à expérimenter.
Ce qui est, en quelque sorte, une attitude plutôt " passive
". Il est, par exemple, certain que le mélèze du Japon,
ou que les douglas américains, proviennent de régions, souvent
fort éloignées du Nord-Est de la France, mais de latitude, donc
de régime photopériodique voisins. L'épicéa commun
de Pologne est, en partie pour la même raison, très intéressant
chez nous. Mais, celui du Nord de la Norvège ne trouve sans doute pas,
en France, le régime photopériodique auquel il est adapté
(jours très longs en été).
On a essayé aussi de " faire monter ", à des épicéas,
quelques degrés de latitude, afin de leur procurer des jours plus longs
en été. On obtient alors souvent une production en volume supérieure,
si les conditions climatiques générales sont favorables, mais
la proportion de bois de printemps (mou) y est plus élevée que
lorsqu'ils se développaient à des latitudes plus faibles.
En ce qui concerne les arbres des régions tropicales, le fait que les
nuits froides diminuent, ou même suppriment la floraison, donc la reproduction
naturelle des espèces de jours courts qui s'y développent habituellement,
permet peut-être d'expliquer qu'elles ne peuvent se maintenir longtemps
dans les stations où les nuits sont normalement assez froides, c'est-à-dire
souvent à partir d'une certaine altitude.
Plus délicates sont les déductions que l'on pourrait tirer de
la modification permanente, dans certains milieux forestiers, de la
qualité de la lumière, dans divers sous-bois. On sait, par
exemple, que les cimes de nombreux arbres feuillus réémettent,
surtout par temps ensoleillé, des rayons infrarouges proches, analogues
à ceux qui provoquent le retour rapide du P. 730 en P. 660, ainsi qu'il
est expliqué plus haut. Or, le P. 730 intervient, en général,
pour inhiber la croissance en longueur, alors que le P. 660 la favorise. On
comprendrait ainsi mieux l'élongation observée, dans certains
sous-bois, chez divers jeunes arbres, feuillus surtout du reste. Cet effet peut,
du reste, s'expliquer aussi par l'intervention, plus classique, de l'ensemble
de la lumière sylvestre sur l'acide ß indole acétique,
qui, lorsqu'elle est élevée, réduit l'élongation,
et lorsqu'elle est faible, permet l'allongement des tiges. Cependant, on comprend
moins bien, par la seule intervention du phytochrome, que les jeunes
arbres résineux soient bien moins longs, dans les sous-bois feuillus,
que ceux qui se développent dans les trouées (9).
D'un autre côté, il paraît difficile de penser au phytochrome,
tout au moins de la façon qui a été indiquée,
pour interpréter les expériences de J. P. NITSCH, rapportées
plus haut. En effet, cet auteur, en apportant au milieu de la nuit un éclairement
supplémentaire blanc ou rouge clair, provoque, vraisemblablement, une
majoration de la proportion du P. 730 (lequel devrait freiner l'élongation).
Or, il obtient, dans certains cas positifs, une majoration très nette
de la longueur des sujets, résineux aussi bien que feuillus. Ce qui semble
contraire aux principes généraux énoncés ci-dessus.
CONCLUSION
Tout ce qui vient d'être dit est donc loin d'être établi
d'une façon définitive, bien que des expériences extrêmement
nombreuses aient été faites, ces dernières années,
pour préciser mieux les divers aspects de ce phénomène.
Les faits, qui servent de base aux théories, sont, selon les expérimentateurs,
souvent discordants. L'ouvrage récent de P. ROLLIN (8), dont la rédaction
est très nuancée, fait bien ressortir la complexité du
phénomène, tel qu'il commence à être envisagé
dans son ensemble. Mais il met aussi en évidence son grand intérêt,
et les multiples applications qui pourront sans doute en résulter, en
agriculture, en horticulture et en sylviculture notamment.
Il ne faut pas oublier, cependant, que certains des processus de la vie des
végétaux ne sont pas, de l'avis à peu près
unanime, considérés actuellement comme rattachés à
l'action du phytochrome (8). On peut citer, dans cet ordre d'idée, le
phototropisme (et les photomorphoses qui peuvent lui être rattachées),
attribué à l'action de certaines radiations - de couleur bleue
principalement - sur l'hétéro-auxine (avec ou sans l'intervention
de substances photo sensibilisantes).
On peut donc, pour résumer, et en simplifiant considérablement,
admettre que la photopériode agit en déclenchant certains rythmes
généraux, chez les végétaux ligneux, comme
la germination, la croissance et la floraison, par exemple, réalisant
les meilleures conditions de leur adaptation aux climats régionaux.
Mais, les " cas particuliers ", fréquents dans le milieu
forestier (végétaux recevant surtout une lumière latérale,
ou commençant leur croissance au milieu d'une végétation
basse très dense), seraient réglés par des réactions
plus localisées, du genre de celles que l'on observe dans l'orientation
phototropique des tiges et des feuillages.
BIBLIOGRAPHIE
(1) DAVID R. (1952). - Les hormones végétales.
Presses Universitaires de France, 187.
(2) GARNER W. W. et ALLARD H. A. (1920). - J. Agr.
Research, 18, 553.
(3) CHOUARD P. (1949). - Expériences de longue
durée sur le photopériodisme ; leçons qui en découlent.
Mémoires de la Société Botanique de France.
(4) MATHON C. C. et STROUN M. (1960). - Lumière
et Floraison. Presses Universitaires de France, 128.
(5) FLINT L. M. et Mc ALISTER E. D. (1937). - Wave lengths
of radiation in the visible spectrum promoting the germination of light
sensitive lettuce seed. Smith. Misc. Collection, 96, 2.
(6) NITSCH J. P. et SOMOGYI L. (1958). - Le photopériodisme
des plantes ligneuses. Ann. Soc. Nation. Hort. Fr., 4e année,
no 16, 466-490.
(7) BORTHWICK H. A., HENDRICKS S. B., TOOLE V. K., TOOLE
E. H. (1951). - A reversible photoreaction controlling seed germination.
Proc. Nat. Acad. Se., 38, 662-666.
(8) ROLLIN P. (1970). - Phytochrome, photomorphogenèse
et photopériodisme. Masson et Cie, 136.
(9) ROUSSEL L. (1972). - Photologie forestière. Masson et Cie, 144.