![]()
Développement des jeunes
arbres en fonction
du rayonnement relatif de leur station (suite)
Cas des arbres feuillus - a) Comportement des chênes rouvre et pédonculé. Ces deux espèces ligneuses ont une importance économique certaine: elles constituent les futaies d'une bonne partie des peuplements forestiers du Nord-Est de notre pays, et se retrouvent, plus au sud, dans certaines régions où elles forment des futaies de haute valeur. Il faut d'abord examiner les conditions de leur installation. Les glands, produits périodiquement (mais non chaque année) par les semenciers, tombent sur le sol en automne. Ils sont recouverts par les feuillages des arbres du peuplement, qui se détachent, à ce moment, des rameaux. Si la fin de l'hiver est favorable (humidité du sol suffisante, absence de froids excessifs), les glands forment, dès ce moment, une radicelle, puis, à partir du mois d'avril, une jeune pousse qui, une fois terminée, se garnit de feuillages. Une seconde pousse (dite de la St Jean) est souvent formée dès le début de juillet.
))
FIG. 40 - Exemples de profils densitométriques (microdensitomètre du C.N.R.F. — POLGE ). Le niveau 4 D correspond à un facteur de transmission optique des écorces de 1/10000. Pour l'explication de la figure, voir le texte ci-dessus.
L'ombre habituelle des couverts forestiers, même les plus denses (de l'ordre de 2 à 3 % de Rr), ne semble guère gêner le déroulement de ces processus (PLAISANCE — 1955, ROUSSEL - 1956 à 1962 ). Ces deux auteurs ont noté un pourcentage de germination élevé, dans toutes les stations (80 %. selon ROUSSEL ). Mais, à partir de la seconde année, les sujets ayant reçu moins de 4 5 % de Rr en été, dépérissent et meurent, sans doute par suite d'une nutrition carbonée nette insuffisante pour couvrir leurs besoins (ROUSSEL - 1956 à 1957 et 1962 à 1963). Ceux qui ont reçu un peu plus de 4 % se maintiennent, très chétifs, pendant quelques années. Par contre, les sujets de plein découvert, à l'âge de 2 ans, sont florissants, feuillés et en parfait état. Les chênes pédonculés sont, en général, plus grands que les chênes rouvres (Fig. 41).
)) |
)) |
FIG. 41 - Chênes rouvres et pédonculés
à la fin de leur seconde année de croissance.
En haut, sous un rayonnement relatif un peu inférieur à 4 %
en été (dépérissement presque total).
En bas, en plein découvert (vigueur satisfaisante) (ROUSSEL 1957).
Dans l'expérience de 1956-1957, 6 caisses de végétation installées du plein découvert (Rr =100 %), jusqu'à un sous-bois de taillis sous-futaie dense (Rr = 3,5 % en été), en passant par 4 états intermédiaires (Rr = 66 %, 57 %, 13,5 % et 4,3%. en été), avaient reçu des glands de chêne rouvre et de chêne pédonculé. Le sol était identique dans toutes les cases. Or, un premier examen
montrait incontestablement que, si le couvert très dense était défavorable, dès la seconde année, pour tous les sujets, le plein découvert, bien que permettant une bonne survie générale, n'était pas, cependant, celui qui donnait les semis les mieux développés. Le tableau de la page suivante, résultant d'un travail très détaillé effectué à l'Institut Botanique de Besançon (TRONCHET. A, TRONCHET. J, GIBOUDEAU & GOGUELY - 1959) est assez démonstratif de l'irrégularité des résultats obtenus.
La figure 42 montre, notamment, que la hauteur des divers sujets de chênes rouvres n'est pas du tout proportionnelle à l'intensité du rayonnement naturel reçu dans chaque station. La même remarque était faite pour les chênes pédonculés. Ceci a paru assez étonnant. En effet, le chêne pédonculé est, comme le chêne rouvre, une " essence de lumière " caractérisée. Or, même pour une " essence d'ombre " résineuse typique (le sapin pectiné par exemple), on n'observe guère que des sujets de 2 ans aient des caractéristiques générales nettement plus favorables à l'ombre qu'à la lumière (ceci sous réserve de l'effet limitant dont il a été parlé plus haut) (Fig. 39).
))
FIG. 42 - Développement des chênes rouvres,
âgés de 2 au, sous des rayonnements relatifs naturels variant de
3,5 à 100 %, en été (GIBOUDEAU 1958).
Essai d'interprétation (ROUSSEL 1958).
Courbe 1 - Action stimulante du rayonnement vertical sur la nutrition
carbonée (photosynthèse).
Courbe 2 - Action freinatrice du rayonnement horizontal sur la multiplication
et l'élongation cellulaires (photoinactivation des auxines).
L'hypothèse " de travail " émise alors fut que, si la composante verticale du rayonnement incident exerce, incontestablement, une action favorable sur la photosynthèse, sa composante horizontale, dont la valeur croît avec l'intensité du rayonnement circumglobal, peut devenir nuisible à la croissance, en filtrant à travers les formations de protection, de la cuticule au phloème, (Fig. 43), et en venant agir, au niveau des cellules en voie de multiplication et d'élongation, comme à celui des vaisseaux transportant l'hétéroauxine libre, ainsi qu'il est dit plus haut (voir page 57). Le phototropisme de ces chênes, se maintenant pendant un grand nombre d'années, constituait une preuve supplémentaire à cette hypothèse.
))
FIG. 43 - Coupe transversale au milieu de la tige épicotylée d'un chêne pédonculé âgé d'un an, développé à l'ombre
| c. = cuticule | cri. = cristaux d'oxalate de calcium |
| ép. = épiderme | Pl = phloème |
| sb. = suber | scl. = scléréides |
| p.c. = parenchyme cortical | f.p. = fibres péricydiques. |
(TRONCHET et GRANDGIRARD 1956).
D'un autre côté, le tableau précédent montre qu'en lumière élevée, l'épaisseur moyenne du suber du chêne pédonculé est plus grande que celle du suber du chêne rouvre (70 à 74 � contre 60 �). Or, le premier, en plein découvert, est plus long que le second (Fig. 41, plein découvert).
))
FIG. 44 - Chênes rouvres à la fin de leur première
année de croissance :
En haut, au centre, sous 1 % de rayonnement relatif en été.
En haut, à droite, en case de végétation à
seul éclairement latéral (lumière à gauche).
En bas, au centre, en plein découvert, à tige non abritée.
A gauche, en plein découvert, mais à tige abritée
progressivement par de petits manchons opaques (ROUSSEL 1957).
 |
FIG. 45 - Chênes pédonculés à la
fin de leur 2e année de croissance. A) Sujets de plein découvert, à tige abritée dès le début de la seconde année. B) Sujets de plein découvert, à tige non abritée et restée en pleine lumière (ROUSSEL 1964). |
 |
FIG. 46 - Chênes pédonculés âgés
de 4 ans, en plein découvert, à tiges non abritées
latéralement (quadrillage 0,15 x 0,15 m) (ROUSSEL 1966). |
Des essais comparatifs de transparence (méthode du papier photographique lent), ont montré, du reste, que cette différence d'épaisseur s'accompagnait d'une densité optique dissemblable. Tout ceci a donc conduit à un essai de dissociation des composantes, verticale et horizontale, du rayonnement circumglobal, dans une station de plein découvert.
Des manchons étroits appliqués sur des chênes rouvres et pédonculés en cours de germination à partir du gland raciné ont montré que les sujets protégés, à 1 et 2 ans, étaient nettement plus longs que ceux qui ne l'étaient pas (s°°) - ROUSSEL - 1957 - (Fig. 44).
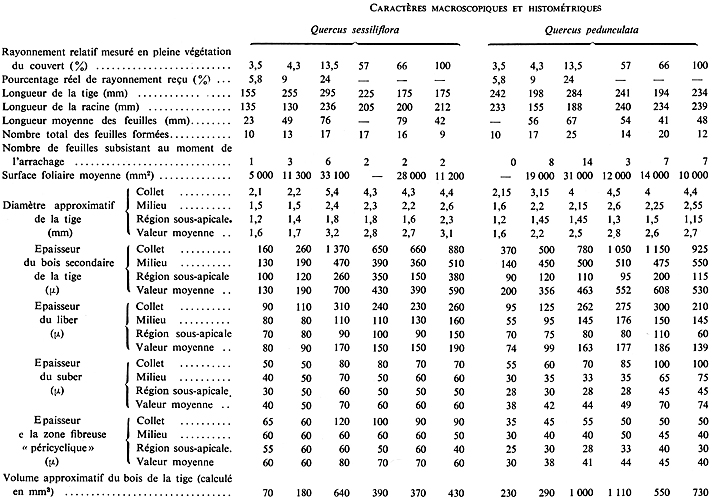
D'autres essais effectués avec des manchons de poterie (0,35 m de hauteur totale) placés autour de certains chênes pédonculés déjà âgés d'une année, ont mis en évidence, dès la seconde année, une élongation nette des sujets protégés par rapport aux témoins (s°°) (Fig. 45). Ces essais, prolongés pendant 3 années (1, puis 2 manchons superposés d'une hauteur de protection d'environ 0,70 m) ont produit, par rapport aux témoins (Fig. 46) des sujets allongés du type représenté à la figure 47. Leurs caractéristiques sont présentées dans le tableau de la page suivante.
La figure 48 représente des microphotographies de coupes faites au C.N.R.F., dans les sens de la longueur, et montrant que, chez les sujets protégés, ce n'est pas la majoration de la longueur individuelle des cellules mais celle de leur nombre qui est responsable du changement de forme constaté. Par ailleurs, des observations faites sur des coupes transversales de tiges, au voisinage du collet, indiquent que le bois du chêne protégé est plus riche en vaisseaux de printemps (mous), et plus pauvre en rayons ligneux (durs), ce qui caractérise souvent un bois de très bonne qualité, facile à travailler.
" L'effet manchon " se traduit également ici, mais à partir de la 3e Ou de la 4e année seulement, par un " transfert des facteurs trophiques de croissance ", comme chez le très jeune épicéa du reste. La tige est longue, mais l'appareil radiculaire est plus réduit chez les chênes protégés (s°°). En somme, on peut penser que, pour un volume total de matière ligneuse fabriquée par un chêne de plein découvert, le rapport : volume tige / volume racine est plus élevé chez le chêne protégé latéralement que chez le chêne dont la tige reçoit la pleine lumière horizontale. Il est à remarquer, du reste, que les expériences qui viennent d'être décrites, exagèrent la dissociation entre les deux composantes, verticale et horizontale, du rayonnement circumglobal naturel, et que, dans le milieu forestier, les différences sont souvent moins marquées.
Quand les chênes vieillissent, en bouquets, dans une trouée pratiquée dans un peuplement plus âgé et dense, ils reçoivent à la fois une lumière verticale convenable, et une lumière horizontale plus réduite. Ils se développent alors d'une façon spéciale et prennent l'aspect classique dit en " cône de régénération " (Fig. 49) qui doit sans doute être attribué à des conditions principalement photologiques.

 |
FIG. 47 - Chêne pédonculé âgé de
4 ans, en plein découvert, à tige abritée à partir
de la seconde année par un, puis par deux manchons de poterie
superposés (quadrillage 0,15 x 0,15)- (ROUSSEL 1966). |
)) |
FIG. 48 - Coupes radiales au milieu de la pousse
formée la
3e année, de chênes pédonculés âgés
de 4 ans, non protégés (témoins) à gauche et
protégés (allongés) à droite (ROUSSEL
1967). |
Les mesures instantanées d'humidité du sol, qui ont été effectuées (PLAISANCE - ROUSSEL) dans de telles formations, donnent, entre le centre et les bords de ces cônes, des différences très faibles, souvent, du reste, inversées (sol plus humide sur les bords qu'au centre). Tout ceci justifie l'intérêt, à côté des considérations de physiologie classiques, de l'intervention de la notion de " coenophysiologie ", de la physiologie de groupe, développée actuellement en U.R.S.S., et dont les conséquences peuvent être intéressantes (YATSENKO. KHMELEVSKI - 1963).

FIG. 49 - Chênes rouvres et pédonculés dans leur 10e été. L'ombrage latéral réciproque des tiges rend très perceptible l'inégalité de répartition de la lumière verticale dans les trouées (formation d'un cône de régénération) (Photo BERNARD).
Cette question semblant à peu près élucidée, quel pourcentage de rayonnement doit recevoir une régénération de chêne rouvre, ou de chêne pédonculé, selon son âge ? Une règle simple consiste à apporter aux jeunes sujets un Rr de 10 %, par année d'âge (10 % dès le début de la première année - 20 % dès le début de la seconde année, 30 %. dès le début de la troisième année ... et le plein découvert à l'âge de 10 ans).
PLAISANCE (1965) a repris cette étude d'une façon systématique, et il conseille, dans la région basse du Jura, 60 à 80 % de Rr en été, pour les chênes pédonculés, à l'âge de 5 ans, et de 30 à 50 % de Rr, en été, pour les chênes rouvres à l'âge de 5 ans également.
Sous réserve que l'alimentation en eau du sol ne soit pas déficiente, on arrive donc très vite, de toutes façons, à la nécessité du plein découvert pour ces deux espèces ligneuses. Mais, en outre, on l'a dit, on doit préférer les traitements qui mettent très tôt les feuillages en lumière, tout en laissant les troncs à l'ombre (ce qui correspond à l'état de futaie régulière).
b) Comportement d'autres espèces feuillues - Seules, quelques études comparables concernant le chêne rouge d'Amérique ont été effectuées, de la façon décrite précédemment, par l'auteur. L'allongement de la tige de cette espèce ligneuse, en abri circulaire latéral, est aussi très net, mais les exigences en lumière verticale (photosynthèse) paraissent plus réduites que pour les chênes rouvre et pédonculé.
Par contre, d'autres espèces feuillues ont été étudiées, d'une façon un peu différente, mais très détaillée, par certains chercheurs étrangers ; on les passera assez brièvement en revue dans les lignes suivantes :
HÊTRE COMMUN - Deux études importantes ont été effectuées récemment sur cette espèce ligneuse intéressante, qui vient sans doute au second rang, après les chênes, dans la production ligneuse de notre pays. La plus ancienne (VEZINA - 1960) se rapporte surtout à des sujets âgés de quelques années. La plus récente, celle de BURSHEL & SCHMALTZ (1965) porte sur de très jeunes sujets. Pour suivre la chronologie du développement du hêtre commun, on commencera par la dernière étude.
Dans 2 stations forestières peu éloignées de Göttingen (Allemagne) l'une à sol de lœss décalcifié, l'autre à sol calcaire, 5 types de caissettes ont été installées, avec 5 répétitions (au total 2 x 5 x 5 = 50 caissettes). La réduction du rayonnement incident était obtenue par des claies ajourées, mais le pourcentage de radiations transmis était mesuré par des appareils enregistreurs. La lumière était principalement distribuée par la partie supérieure des caissettes. Ces dispositifs, comme il a été dit à la page ci-dessus, donnaient des rayonnements relatifs de 100 % - 77 % - 19 % - 12 % et 1 %. Ils ont reçu au printemps 1962 des semis de hêtre âgés de 1 an, pour moitié d'origine naturelle, Prélevés en forêt, et pour moitié extraits dans une pépinière. Les résultats furent obtenus au bout d'un an (automne 1962) et de deux ans (automne 1963). Voici quel fut, dans chaque type de sol, et pour chaque sorte de semis, la production de matières sèches obtenue au bout de 2 ans, exprimée en grammes par mètre carré :
|
Rayonnement relatif
|
Sol de loess
|
Sol calcaire
|
||
|
Plants de pépinière
|
Plants naturels
|
Plants de pépinière
|
Plants naturels
|
|
|
100%
|
748
|
365
|
823
|
302
|
|
77%
|
553
|
298
|
816
|
277
|
|
18%
|
366
|
158
|
435
|
141
|
|
12%
|
195
|
90
|
269
|
77
|
|
1%
|
26
|
7
|
53
|
15
|
Ayant analysé les variations des principaux facteurs physiques des stations, les auteurs ont effectué des calculs de tests statistiques qui démontrent que c'est, de beaucoup, le facteur " intensité du rayonnement naturel " qui a exercé la plus grande influence sur la croissance des jeunes plants (sauf à partir de 77 % de Rr sur sol calcaire). Les autres facteurs interviennent assez peu, ou même pas du tout.
Par contre, la croissance en hauteur est restée assez stable, tant que le Rr est resté compris entre 20 % et 100 %. Du point de vue de la composition chimique des jeunes hêtres, à l'ombre le pourcentage de matières minérales incorporées dans le petit arbre est relativement plus important, qu'en lumière, où le pourcentage des polyholosides prédomine, ceci en relation évidente avec l'intensité plus grande de la photosynthèse. En résumé, les sujets naturels manifestent, à 2 ans, un accroissement de poids de 70 à 150 kg par hectare dans les stations sombres, et de 3000 kg par hectare dans les stations les mieux éclairées. La différence est, on le voit, très sensible.
HÊTRE COMMUN ET FRÊNE COMMUN -VEZINA (1960) a réalisé un très important travail sur la répartition du rayonnement naturel, ou de la seule lumière, et des précipitations reçues au sol dans diverses stations forestières. Il a effectué simultanément des observations sur l'influence de ces facteurs physiques sur la croissance de fourrés naturels de hêtre commun et de frêne commun, dans des forêts des environs de Zurich. L'âge des sujets était voisin de 10 ans. Ont été étudiées les caractéristiques suivantes :
Nombre des tiges : Quand le Rr passe de 29 % à 58 %, le nombre des tiges de hêtre passe de 1425 à 1731 - quand le Rr passe de 23 % à 55 %, le nombre des tiges de frêne passe de 230 à 731 (par hectare).
Composition des fourrés : La valeur du Rr a peu d'influence.
Structure selon les classes de hauteur et de vitalité : Quand le Rr augmente, on constate un déplacement de la courbe de répartition des tiges vers les classes supérieures de hauteur. Les sujets ont également une vitalité plus grande.
Longueur de la pousse annuelle des arbres dominants : Pour le hêtre, elle croît modérément avec l'accroissement de la valeur du Rr : sous 8 %, longueur de 17 cm, sous 58 %, longueur de 29 cm. Ceci en assez bon accord avec les observations de BURSHEL & SCHMALTZ. Pour le frêne, les différences sont bien plus marquées : sous 7 %, longueur de 5 em, sous11 % longueur de 8 cm, sous 23 %, longueur de 55 cm et sous 53 %, longueur de 77 cm.
Mortalité due aux gelées printanières : Elle croît quand le découvert (donc le rayonnement nocturne) augmente.
En conclusion, VEZINA estime qu'un Rr de 50 % est le plus convenable pour le frêne vers l'âge de 10 ans - un peu moins pour le hêtre, sans qu'une valeur chiffrée soit donnée.
Cet auteur a effectué, en serre, diverses expériences sur des semis de frêne. Dans l'essai le plus complet (1958), 4 éclairements relatifs (85 % -50 % -22 % et 12 %), 2 degrés d'humidité du sol et 2 types de terres (sol brun et sable) ont été employés. Le dispositif était répété 6 fois (96 vases d'expériences au total).
Dans tous les cas, c'est le Rr (plus exactement l'Er) le plus fort qui a donné les meilleurs résultats (poids moyen de tige sèche le plus élevé). En moyenne, les chiffres obtenus sur des sujets âgés de 3 ans, au bout de 5 mois d'observation, sont les suivants :
|
Éclairement relatif
|
Gain sur le poids sec de tige
(gr)
|
Gain sur la hauteur de tige (cm)
|
|
85%
|
115,6
|
17,25
|
|
50%
|
91,6
|
15,55
|
|
22%
|
67-
|
14,75
|
|
12%
|
56,7
|
13,20
|
Les tests statistiques donnent une liaison extrêmement probable entre le poids sec ainsi que la hauteur des tiges et l'intensité de la lumière (s°°). Ce sont surtout les sols riches, et bien arrosés, qui favorisent le mieux l'utilisation optimale du rayonnement naturel. Mais, dans chaque type d'éclairement, ce sont surtout les sols les plus humides qui se sont révélés les plus favorables. En somme, on trouve ici la confirmation que rayonnement et humidité du sol sont indispensables à la croissance du hêtre et du frêne, comme de tous les végétaux du reste ...
Essences d'ombre et essences de lumière - Une notion classique en sylviculture est celle des " essences d'ombre " et des " essences de lumière " ; on a beaucoup écrit sur cette question, chaque sylviculteur pense détenir le sens exact qu'il faut lui attribuer, en oubliant de remarquer que, dans la quasi-totalité des cas, il n'a jamais employé des procédés, ou des instruments, même rudimentaires, qui auraient contribué à fortifier, ou bien à modifier sa position. Car, et c'est là le fond même du problème, on discute, scolastiquement, de problèmes qui ne peuvent que recevoir des solutions physiologiques.
Dans un Traité relativement récent, PERRIN (1952) donne la classification suivante, telle qu'elle était admise par l'Allemand GAYER, en 1898 :

D'après les résultats obtenus par les mesures précises qui viennent d'être rapportées, on verra que cette classification appelle certaines réserves, et des corrections assez sérieuses.
DE LIOCOURT & SCHAEFFER (1968) ont récemment insisté sur les incertitudes que présentent des classifications de ce genre (car il y en a eu bien d'autres), et sur l'intervention probable d'autres facteurs que la lumière, en cette matière toujours controversée.
Il est évident d'abord, on l'a dit, que les premiers sylviculteurs qui, d'une façon assez générale du reste, ont admis cette notion, n'ont jamais, et pour cause, effectué de mesures précises de lumière, aucun appareil satisfaisant n'étant alors à leur disposition. Mais, ils ont constaté que, dans des trouées, recevant visiblement une assez forte quantité de lumière, certaines espèces s'étaient installées naturellement et avaient persisté ; que par contre, dans des sous-bois denses, où la lumière était manifestement réduite, d'autres espèces s'étaient installées et s'y étaient maintenues. Ils n'ont pas réalisé exactement cette sorte de balancement entre les facteurs énergétiques et hydriques, en particulier, et qui n'a été mis en évidence, dans de nombreuses régions, que récemment (voir page 73).
Leurs observations étaient sans doute pertinentes (en fait, leur sens forestier était bien plus développé que celui de beaucoup de nos modernes sylviculteurs) - mais leur interprétation était incomplète.
On pourrait penser que l'activité photosynthétique unitaire des différentes variétés de chaque espèce ligneuse apporte des éléments chiffrés en faveur de la classification habituelle. Or si, incontestablement, le mélèze d'Europe, par exemple, ou bien le chêne pubescent, ont une assimilation carbonée plus intense, en conditions de lumière égale, que le sapin pectiné, l'épicéa ou même que le chêne vert, cette différence correspond, plutôt, au fait très apparent, que les premières espèces ne conservent leurs appareils foliacés que pendant quelques mois, alors que les secondes ont la possibilité de les faire fonctionner pendant toute l'année (Fig. 25). Par contre, pour les résineux, d'autres types de relations viennent corroborer l'idée que certaines espèces ligneuses utilisent mieux la lumière, même réduite, des sous-bois que d'autres. Ainsi, comparant l'activité physiologique du sapin pectiné, de l'épicéa commun et du pin sylvestre, on trouve, dans l'étude citée de LARCHER (1969) :
- que le " point de compensation " (éclairement le plus bas au-dessous duquel la respiration des aiguilles réutilise plus de matières hydrocarbonées que la photosynthèse n'en fournit), est, pour les 3 espèces ci-dessus, de plus en plus élevé: respectivement 300 lux, 400 lux et 1000 lux ;
- que " l'éclairement de saturation " (au-dessus duquel la photosynthèse nette n'est plus sensiblement majorée), varie dans le même sens, soit respectivement un peu plus de 20 000 lux, un peu plus de 30 000 lux et environ 60 000 lux.
Ceci correspond, dans ses grandes lignes, très partiellement à la classification ci-dessus de GAYER (l'épicéa serait plutôt une essence de demi-lumière) et explique les résultats globaux des observations effectuées dans le milieu forestier (voir pages 82 à 98).
Pour les arbres feuillus, une relation un peu décevante pour les forestiers physiologistes est établie par LARCHER (1961-1969) et par RETTER (1965): le hêtre commun (d'altitude modérée dans la région d'Innsbruck) et le chêne vert (dans la région du Lac de Garde) ont, l'un et l'autre, 2 sortes de feuilles, d'ombre et de lumière, qui réagissent dans chacun de ces deux cas, à peu près de la même façon, à une majoration de l'éclairement (Fig. 50). En somme, le hêtre, considéré comme une " essence d'ombre ", aussi bien que le chêne vert, considéré comme une " essence de demi-lumière " ont des feuilles qui, selon leur situation dans les cimes, se comportent, soit comme celles des plantes d'ombre, soit comme celles des plantes de lumière, ainsi que LUNDEGARDH l'avait établi, dès l'année 1921. Il n'y a donc aucune possibilité d'utiliser ces résultats pour tenter de différencier ces deux espèces ligneuses.
))
FIG. 50 - Absorption du gaz carbonique en fonction de l'éclairement, pour les feuilles d'ombre et de soleil du hêtre commun et du chêne vert,au voisinage de l'optimum de température (LARCHER 1961 -1969, RETTER - 1965).
Toutefois, il semble que la considération du " point de compensation " permette de comprendre pourquoi le hêtre commun (minimum de 200 à 300 lux) sera un peu favorisé, dans les sous-bois denses, par rapport au chêne vert (minimum de l'ordre de 500 à 600 lux).
JACQUIOT (1970) estime pouvoir, de cette façon, expliquer certains phénomènes d'installation successive, dans des peuplements évoluant à la suite d'un incendie (bouleaux verruqueux à couvert léger permettant l'installation des pins sylvestres, auxquels succèdent des chênes, rouvres et pédonculés, favorisant, en définitive, la constitution de semis de hêtres communs).
Un autre genre de processus est à signaler, dans le cas des " essences d'ombre " et des " essences de lumière " : il concerne l'alimentation des racines en eau et en matières minérales diverses, en fonction des espèces en cause et de la station où elles se développent. On a déjà vu (page 106 ci-dessus) que les chênes, rouvres et pédonculés, abrités latéralement (ou se développant à l'ombre dense) ont une tige allongée, et, corrélativement, au bout de quelques années, un appareil radiculaire plus réduit que celui des chênes en permanence bien éclairés. Le même genre de phénomène se constate, mais précocement, pour l'épicéa commun (Fig. 29). Une expérience récente, non publiée (ROUSSEL - 1969), portant sur 6 espèces résineuses, abritées et non abritées, permet de dresser le petit tableau qui suit, concernant les rapports de la longueur moyenne de l'axe hypocotylé (A), à la longueur moyenne de la radicelle (R) - (s°) :
p 116
|
Groupes
|
Espèces en expérience
|
État des semis à
l'âge de 3 mois
(Rapports A/R) |
||
|
Témoins non
abrités |
Témoins abrités
(abri D = H) |
|||
|
I
|
{
|
Mélèze des Alpes |
1,56
|
1,93
|
| Pin sylvestre des Vosges |
1,11
|
1,47
|
||
|
II
|
{
|
Épicéa du Jura |
0,68
|
1,12
|
| Épicéa des Alpes du Sud |
0,67
|
1,17
|
||
|
III
|
{
|
Sapin des Vosges |
0,74
|
0,94
|
| Sapin de l'Aude |
0,79
|
0,69
|
||
Pour les sujets du groupe I (essences dites de pleine lumière) les axes sont toujours nettement plus longs que les radicelles - pour les sujets du groupe III (le sapin des Vosges est une essence d'ombre typique, le cas du sapin de l'Aude est plus incertain), les radicelles sont toujours plus longues que les axes. Pour les sujets du groupe II (essences d'ombre, selon GAYER - essences " tolérantes " selon plusieurs praticiens), les radicelles sont Plus longues que les axes à la lumière, et les axes sont plus longs que les radicelles à l'ombre assez dense. Il y aurait peut-être, dans cette classification, un élément susceptible d'expliquer l'installation et le maintien, sous le couvert, du sapin des Vosges tout au moins (espace de sol prospecté relativement important), la disparition plus ou moins rapide, du mélèze d'Europe et même des pins sylvestres sous un couvert dense (espace de sol prospecté plus réduit qu'en pleine lumière) - et la relative plasticité de l'épicéa commun, " essence ondoyante et diverse ", pour prendre les termes de l'un de nos auteurs classiques. De toutes façons, ce type de raisonnement fait intervenir, non plus le seul rayonnement naturel, mais le complexe : rayonnement / eau, qui varie dans chaque station forestière, et dont il est difficile de séparer les effets, à la période véritablement cruciale de l'installation des semis naturels.
En somme, et pour résumer ce qui vient d'être dit précédemment, on pourrait admettre, tout au moins à titre provisoire :
1° que les " essences d'ombre " s'installent assez bien sous le couvert, parce qu'elles ont des racines qui restent assez longues au moment de leur germination, permettant leur bonne alimentation en eau, et aussi, parce que leur " point de compensation " (lumière relative pour laquelle la photosynthèse brute égale la respiration) est situé sous un éclairement assez bas: 2 à 300 lux pour le hêtre commun, et le sapin pectiné. Par la suite, elles profitent d'une majoration -modérée de l'Er (jusqu'à environ 25 % pour le sapin pectiné), alors qu'au-dessus, leur croissance, tout au moins en hauteur, ne s'améliore guère. La saturation lumineuse serait assez vite atteinte.
2° que les " essences de lumière " se rencontrent plus rarement sous le couvert, en raison de l'élongation de leurs tiges, corrélative d'un raccourcissement de leurs racines (alimentation en eau moins favorable), et parce que leur point de compensation est plus élevé: 5 à 600 lux pour le chêne pubescent, 1000 lux pour le pin sylvestre. Elles profitent d'un Er plus fort (jusqu'à 50 % pour l'épicéa, et jusqu'à 80 ou 100 %. pour le pin sylvestre et le mélèze). Les chênes, rouvre et pédonculé, ont, la première année, un point de compensation à peine plus élevé que celui du hêtre, mais il s'élève par la suite. La saturation lumineuse ne serait atteinte, pour les " essences de lumière " âgées de quelques années, que pour 30, 40 ou 50000 lux, soit à peu près, ou même davantage que la moyenne du plein découvert (voir page 38).
Remarques sur le comportement général des résineux et des feuillus - A la page 90 on a montré que les très jeunes résineux sont phototropiques, s'allongent en abri latéral circulaire (effet manchon), puis perdent rapidement ces facultés ; leur développement général est alors intimement lié à l'intensité du rayonnement naturel qui atteint leurs aiguilles.
À la page 106 on a montré par contre que les arbres feuillus, ont, dans leur jeune âge, un développement général lié à leur nutrition carbonée, mais qu'ils peuvent être freinés dans leur croissance par une intensité trop élevée du rayonnement à direction horizontale (phototropisme, élongation en abri latéral circulaire), et que cet effet se poursuit pendant un grand nombre d'années. La figure 51, extraite d'un travail déjà ancien d'ENGLER (1924) est tout à fait démonstrative de cette différence de réaction, pour des sujets âgés de 20 à 30 ans au moins. Le résineux (un épicéa commun) reste bien rectiligne - le feuillu (un bouleau verruqueux) continue à s'orienter vers la lumière. Du reste, ENGLER a obtenu la déformation et même la rupture de tuteurs robustes placés contre de jeunes arbres feuillus (des frênes communs) pour tenter de maintenir leur croissance verticale, en conditions de lumière déséquilibrée.
L'effet ralentisseur de la lumière sur la croissance se maintient chez les feuillus, alors qu'il est fugace chez les résineux.
Or, on admet en général, que les gymnospermes, et en particulier que les coniférales, auxquelles appartiennent toutes les espèces résineuses étudiées, ont commencé à apparaître vers la fin de l'ère primaire (au permien) et qu'elles étaient en plein développement au secondaire, pour régresser légèrement depuis cette époque - alors que les angiospermes, auxquelles se rattachent les espèces feuillues mentionnées, ne se sont manifestées qu'à la fin de l'ère secondaire (au crétacé) et n'atteignent qu'actuellement leur plein développement (DEYSSON -1961).
Doit-on alors considérer que l'on se trouve en matière d'évolution, en présence de l'un de ces cas assez curieux où, selon la définition qu'en donne
le biologiste J. ROSTAND, " ce sont les caractères de l'embryon, ou du jeune, donc transitoires chez l'ancêtre - qui persisteront chez le descendant jusqu'à l'âge adulte, pour s'intégrer au type normal " ? On a proposé, pour ce type de phénomène évolutif, la dénomination de " coenogénèse prophétique " (OSTOYA - 1951).
))
Pour certains autres caractères des conifèrales, GAUSSEN (1937-1945) a déjà soutenu cette idée quand il a fait remarquer que " dans une espèce, un caractère susceptible d'évolution peut apparaître plus développé chez le jeune que chez l'adulte ". Dans ce cas, selon cet auteur " le jeune indique donc le sens de l'évolution future".
En ce qui concerne l'aptitude phototropique, fugace chez les coniférales, et persistante chez les fagales, la question vaut, en tous cas, la peine d'être posée ...
![]()