Cas des arbres résineux -
La dénomination exacte des arbres étudiés est donnée
à la page 18
ci-dessus. Il est à remarquer que chaque espèce (et cette remarque
semble spécialement valable pour l'épicéa commun), comprend
de nombreuses variétés locales, considérées comme
stabilisées, et dont le comportement peut être très différent.
Chaque fois que ceci sera possible, on précisera la provenance du végétal,
et la région où il a été expérimenté.
Une aide très précieuse a été apportée à
l'auteur par J. BOUVAREL, Directeur de Recherches au C.N.R.F. de Nancy, qui
lui a fourni de nombreux lots de graines d'origine contrôlée.
L'auteur, depuis 25 années, a étudié
systématiquement le comportement d'espèces résineuses variées,
la plupart du temps en cases de végétation du type décrit
plus haut :
- Dans le Haut Jura (altitude de 800 à 1000 m environ),
sur un sol de rendzine calcaire plus ou moins profonde, et sous des précipitations
moyennes annuelles abondantes (de 1200 à 1500 ou 1800 mm par an), la
base de référence a été l'épicéa commun,
variété du Haut Jura - puis a été étudié
le sapin pectiné du Haut Jura, puis 4 autres espèces indigènes,
puis 6 espèces non indigènes.
- En Champagne humide (altitude 150 m environ) sur un sol
profond et frais de limon des plateaux, et sous des précipitations modérées
(de 600 à 800 mm par an), la base de référence a été
également l'épicéa commun, variété du Haut
Jura - et, à côté, ont été étudiés
11 autres résineux susceptibles d'être utilisés dans les
reboisements français.
a) Epicéa commun. - L'épicéa commun du Jura germe aussi bien (taux de 60 à 70%) sous le rayonnement relatif réduit des sous-bois denses (de l'ordre de 2 à 3 %), qu'en pleine lumière. A ce moment, utilisant les réserves de sa graine (germination épigée), il réagit à la lumière latérale et manifeste un phototropisme très net, mais temporaire (Fig. 28) (s°°). Pendant cette période de phototropisme juvénile, il est également sensible à la réduction de la lumière latérale circulaire, qui provoque chez lui, par suite sans doute d'un " transfert des facteurs trophiques de croissance " (CHAMPAGNAT), un allongement de son axe hypocotylé (du collet au niveau des premières aiguilles), et un raccourcissement corrélatif de sa radicelle (Fig. 29) (s°°).
))
FIG. 28 - Phototropisme juvénile
de l'épicéa commun du Jura, au cours de son 1er été
(quadrillage 0,01 x 0,01 m) (ROUSSEL 1963).
Cette simple modification de forme, qui se rencontre du reste chez les très jeunes épicéas germant, en pleine forêt, sous les couverts denses, explique en partie la disparition rapide, lors d'un dessèchement superficiel du sol, des sujets d'ombre, d'apparence pourtant très florissante, et que l'on enregistre parfois, au cours des étés secs suivant une bonne année de semence, dans le Haut Jura. Les sujets ayant germé en lumière, à axes plus courts, mais à racines plus longues, résistent mieux à cette sécheresse superficielle. Au surplus, les appareils foliacés deviennent, très vite, plus développés (Fig. 36). On peut comprendre, peut-être, de cette façon l'utilité de la " coupe d'ensemencement ", pratiquée par certains sylviculteurs pour cette espèce ligneuse, et qui doit précéder la chute des graines au sol.

FIG. 29 - Modification de la forme de
l'épicéa commun du Jura, suivant la hauteur de l'abri latéral,
au cours de son 1er été.
A gauche, sujet de plein découvert, à
radicelle longue.
En allant vers la droite, sujets protégés
de la lumière latérale par des abris de plus en plus hauts (jusqu'à
0,05 m) à radicelles de plus en plus courtes. (ROUSSEL 1969).
Il est à noter que pour d'autres variétés d'épicéa
commun, celle des Alpes du Nord en particulier, ce type de réaction est
moins régulier, et ceci explique peut-être certaines observations
faites sur les conditions spéciales de leur installation (s°°).
Le phototropisme de l'épicéa commun du Jura se manifeste, en case de végétation à seul éclairement latéral (le rapport des éclairements reçus sur les faces internes et externes des tiges est voisin de 1/20), pendant la seconde année de la croissance, mais plus faiblement. Dans les mêmes cases de végétation il disparaît dès le début de la troisième année et la pousse du jeune épicéa, même en lumière déséquilibrée, reste complètement rectiligne (Fig. 30)

FIG. 30 - Dans le cours de son 3e été, l'épicéa commun du Jura, en case de végétation à seul éclairement latéral (la lumière venant de la droite) se redresse - d'une façon un peu exagérée (au bas de la figure) — mais il est bien moins développé que son homologue développé entièrement en plein découvert (en haut de la figure). Le quadrillage est de 0,15 x 0,15 m (ROUSSEL 1965).
 |
 |
FIG. 31 - Epicéas de 7 ans situés
sous un Rr variable de 5 à 50% (milieu naturel)
(ROUSSEL 1949).
(s°°). Du reste, comme la lumière
générale est plus faible qu'en plein découvert, dès
la fin de sa première année, et surtout au cours de ses seconde
et troisième années, la pousse annuelle du sujet bien éclairé,
réalisant une photosynthèse satisfaisante, est plus épaisse
et plus longue que celle du sujet situé dans la case de végétation
à seul éclairement unilatéral. Des observations analogues
sont faites, en permanence dans le milieu forestier, sur l'absence de phototropisme
de l'épicéa commun, dès sa seconde ou sa troisième
année.
Il est à noter que si, sur des sujets
âgés de 4 ans devenus, comme il est dit, non phototropiques, on
tente, par un abri latéral, généralisé (large manchon
de poterie placé autour du sujet) - ou localisé à la seule
pousse annuelle (tube placé à l'extrémité d'une
tige fichée en terre), de réduire la lumière latérale
qui atteint la pousse en voie de développement on n'obtient plus aucun
effet d'élongation (s°°). Il y a donc, semble-t-il, une liaison
entre la faculté phototropique (de durée assez courte), et la
réaction d'allongement à l'abri latéral.
Par la suite, on constate que, dans le milieu naturel, il existe une corrélation très nette entre le développement général du sujet, du diamètre et de la longueur de sa pousse annuelle, ainsi qu'entre l'importance de son appareil radiculaire - et le rayonnement relatif qui règne dans la station (Fig. 31 et 32). Quand le rayonnement relatif atteint 50 % environ, l'influence heureuse des radiations s'affaiblit, et il semble que, jusqu'à l'âge de 20 à 25 ans, d'une façon approximative, il n'y ait plus grand intérêt à trop dépasser cette valeur. Cette notion peut être économiquement utile, dans la conduite des opérations de dégagement des plantations, ou des régénérations naturelles, car ces travaux coûtent d'autant plus cher qu'ils sont plus poussés. Quand les arbres deviennent plus âgés, on n'a pas de données bien précises sur leurs exigences en radiations, mais on pense, généralement, que c'est le plein découvert qui convient le mieux, sous réserve que l'alimentation en eau du sol soit convenablement assurée.
))
FIG. 32 - Corrélations observées
entre le rayonnement ou l'éclairement relatif du sous-bois de diverses
sapinières et pessières du Jura, et certaines caractéristiques
des sujets d'épicéa commun qui s'y développent naturellement
(ROUSSEL 1952)
I - Hauteur totale du plant de 4 ans (1943-1946). II - Hauteur totale du plant
de 7 ans (1943-1949). III - Longueur de la pousse 1948 de plants de 15 à
20 ans (Levier). IV - Longueur de la pousse 1951 de plants de 15 à 25
ans (Gd-Côte).
On peut se poser la question de savoir, pour
quelles raisons physiologiques, les épicéas communs du Jura (comme
du reste un très grand nombre d'autres résineux), après
s'être montrés sensibles à la lumière latérale,
deviennent, au bout de 2 à 3 ans, insensibles à cette influence.
L'auteur a émis l'hypothèse que la densité optique (D)
des formations de protection (pratiquement des écorces, depuis l'épiderme
jusqu'au cambium) pouvait jouer un rôle dans le déroulement des
divers processus décrits. Cette densité optique peut être
mise en évidence par simple application sur un papier photographique
lent (méthode des photogrammes), ou par mesure directe dans un microdensitomètre
(appareil utilisé par POLGE au C.N.R.F. de Nancy).
En général très faible
chez le jeune sujet (D voisin de 2, soit un pourcentage de transmission
optique de 1/100), elle est fortement majorée quand le petit arbre prend
de la vigueur (D passe alors à 3 ou 4, soit un pourcentage de transmission
optique de 1/1000 à 1/10 000, parfois moins). La lumière horizontale
qui atteint l'ensemble des cellules de cet arbre est faible, mais non négligeable,
lorsqu'il est très jeune (25 à 50 lux la première année,
par moments), mais diminue rapidement (5 lux, puis 0,5 lux au cours des 2e
et 3e années). Un effet ralentisseur peut donc être
observé dans les premiers cas, par action d'inhibition partielle sur
l'hétéroauxine, au moment de son transfert, ou bien au niveau
des cellules mêmes, où elle exerce habituellement son action. Un
freinage de l'activité desdites cellules peut également être
invoqué (page 57
ci-dessus). Cette hypothèse du rôle de la densité optique
des formations de protection se retrouve, du reste, à propos des autres
résineux, et également des jeunes chênes (voir page
106 ). On pourra voir, à la figure
40, comment se présente un profil densitométrique, tel qu'il
est obtenu avec le microdensitomètre dont il a été question
plus haut. On y remarque la forte différence relevée entre la
densité optique des branches d'épicéa commun, dont la direction
n'est pas modifiée par la lumière, et celle des branches de pin
sylvestre, qui, à l'intérieur des massifs, se retournent
nettement vers la lumière extérieure.
Il est à remarquer enfin que l'on peut, d'une façon artificielle, provoquer un léger retour au phototropisme juvénile, chez quelques résineux, dont l'épicéa commun du Jura, en les plaçant dans des cases de végétation spéciales sous une lumière réduite et fortement déséquilibrée (le rapport entre les éclairements internes et externes des tiges étant passé à 1/50 environ).
A 6 ans, par exemple, l'épicéa
commun manifeste alors une légère orientation de sa pousse principale
vers la lumière (10° environ) ; mais, dans cet éclairement
général faible, la densité optique des écorces est
revenue à 2 ou 3 environ (au lieu de 4 et 5 pour les sujets de pleine
lumière), et, de toutes façons, un tel microclimat ne se rencontre
pratiquement pas dans la pratique sylvicole usuelle.
b) Sapin pectiné - Les variétés
de sapin pectiné étudiées (provenance : Jura - Vosges
- Alpes - Aude) se comportent d'une façon assez semblable à celle
qui a été décrite plus haut, dans leur première
jeunesse.
Comme l'épicéa commun, dès
leur germination (en général moins satisfaisante, à l'ombre
comme à la lumière - le taux de germination étant souvent
de l'ordre de 20 à 25 %), les jeunes sapins pectinés sont phototropiques,
mais d'une façon peut être moins marquée. Dans le milieu
naturel, en lumière déséquilibrée (régénérations
de lisière) on observe souvent une forme en S. L'abri latéral
circulaire favorise l'élongation des axes hypocotylés et le raccourcissement
de la radicelle (s°°). Mais celle-ci demeure relativement longue,
même quand le sujet s'est développé dans une ombre dense.
Il est à remarquer que le développement du sapin pectiné,
à l'âge de 2 ans, n'est pas très différent, que le
sujet ait poussé sous le couvert ou en pleine lumière (Fig. 39).
Le phototropisme juvénile du sapin pectiné
s'atténue au cours de la seconde année ; à la fin
de celle-ci, on a observé, chez celui des Vosges, la formation, du côté
ombragé, d'un bourgeon qui, dès le début de la troisième
année, donne naissance à une pousse bien verticale, et non phototropique.
La densité optique des formations de protection de cette pousse est plus
élevée (D = 3 à 4), que celle de la pousse primitive,
restée phototropique et se prolongeant en rameau latéral (D
= 2 à 3). En case de végétation à seul éclairement
latéral, la poursuite de l'expérience montre qu'à partir
de la troisième année, la pousse principale et les rameaux développés
du côté ombragé ne sont plus sensibles à la lumière
latérale. Par contre, les aiguilles continuent pendant plusieurs années
à s'orienter perpendiculairement à cette lumière (réaction
dite de " parahéliotropisme "). À partir de 4 ans, le
sapin pectiné reste insensible à l'abri circulaire latéral
(s°°). Là encore, on remarque cette évolution parallèle
entre la perte de la faculté phototropique, et l'absence de réaction
à l'ombragement de la tige.
Au cours des essais effectués dans le
Jura, le sapin pectiné a supporté tous les degrés d'éclairement,
de l'ombre dense au plein découvert. Par contre, en Champagne, au cours
d'un été très chaud et très sec (1964) à
partir d'un rayonnement relatif de 35 % en été, les variétés
du Jura et des Vosges ont à peu près disparu, alors que la variété
de l'Aude se maintenait parfaitement.
A l'âge de 15 à 20 ans, dans le Jura, le besoin en rayonnement relatif du sapin pectiné du Jura est satisfait jusqu'à une valeur d'environ 25 %.
Au-dessus, on n'a pas constaté de croissance plus favorable (figure 33). Cette considération est importante car elle marque les réserves avec lesquelles on doit accueillir la dénomination " d'essence d'ombre " qui est, en général, appliquée à cette espèce ligneuse. Le sapin pectiné supporte l'ombre, mais il bénéficie, cependant, d'un accroissement de lumière jusqu'à une valeur relativement élevée. BLUTEL (1969), à l'occasion d'observations effectuées sur des plantations de sapins pectinés sous des taillis pauvres en réserves de Franche-Comté, région basse, formule des conclusions absolument analogues.
))
FIG. 33 - Corrélations observées
entre le rayonnement relatif dans le sous-bois (Rr) d'une sapinière,
et la longueur de la pousse annuelle de sujet d'épicéas communs
et de sapins pectinés, âgés de 10 à 15 ans et installés
naturellement.
Les résultats sont présentés sous forme de courbes
de fréquence (ROUSSEL 1954)
On a peu de données sur les exigences
en lumière, vers l'âge de 15 ou 20 ans, du sapin pectiné
de l'Aude ; dans les régions de plaine où il est habituellement
introduit, il paraît plus résistant aux chaleurs estivales, mais
réagirait au couvert de la même façon que son homologue
du Jura.
En ce qui concerne la variété
de sapin pectiné, que l'on rencontre en Italie centrale (région
des Apennins), GIACOBBE (1969) a observé qu'une insolation assez intense
était, non seulement bien tolérée, mais aussi très
utile, par l'effet de réduction des phénomènes de transpiration
au niveau de la tige qu'elle provoque, à la bonne installation et au
premier développement de cette espèce. (Voir page
69).
Il est enfin probable que l'arbre adulte demande,
si l'alimentation en eau du sol est satisfaisante (conditions remplies dans
les régions montagneuses de l'Est de la France), la totalité du
rayonnement reçu par une station de plein découvert.
c) Autres espèces ligneuses. -
Les réactions aux variations de l'intensité du rayonnement naturel
ont été également étudiées chez de nombreuses
autres espèces ligneuses. En fait, les observations ont porté,
à un degré plus ou moins avancé, sur presque toutes les
sortes de résineux énumérées à la page
18 ci-dessus, et il serait fastidieux, pour le lecteur, de les rapporter
en détail.
On peut d'abord préciser que ces espèces
se comportent, dans leur toute première jeunesse, à peu près
de la même façon que l'épicéa commun et que le sapin
pectiné : pourcentage de germination, chez chaque espèce,
comparable dans les sous-bois denses et en plein découvert - phototropisme
juvénile très net en cases de végétation à
éclairement unilatéral et allongement corrélatif de l'axe
hypocotylé, en abri circulaire latéral. Puis, vers la fin de la
seconde année, et au cours de la troisième année de développement,
réduction très sensible de ces facultés, dans les conditions
habituelles des microclimats forestiers. Par contre, dépendance assez
étroite entre la croissance et l'intensité du rayonnement naturel
circumglobal.
Cependant comme pour l'épicéa
commun du Jura, il est possible, en plaçant les sujets dans des conditions
de souffrance et en lumière fortement déséquilibrée,
d'obtenir un léger retour au phototropisme juvénile de la majorité
des espèces qui ont été expérimentées - à
l'exception des sapins. Dans le cas du pin sylvestre, race de la Champagne humide,
le retour au phototropisme est relativement facile à obtenir, même
au bout de 10 à 15 ans, mais cet effet, très marqué pendant
un ou deux ans, se termine en général par le dépérissement
complet des sujets (ROUSSEL - 1967).
Une observation que l'on peut faire fréquemment, dans le cas du pin sylvestre, est le phototropisme des branches développées sur des arbres
de lisière, du côté de
l'intérieur du massif et qui se retournent alors, dans un plan presque
horizontal, vers la lumière. Dans les mêmes conditions, les branches
intérieures des épicéas de même âge, en même
type de sol, ne manifestent aucun phototropisme. Ces différences de réaction
s'accompagnent, on l'a dit, d'une façon très nette, d'un changement
dans la densité optique des formations de protection (Fig. 40).
La figure 34 représente l'état, à un an, de 6 espèces résineuses indigènes développées dans des cases de végétation situées en pleine forêt, sous 4 rayonnements relatifs différents, dans des massifs des hautes chaînes du Jura (ROUSSEL - 1953). Leur reproduction sur papier millimétrique permet de comparer leur comportement sous 6 %, 12 %, 37 % et 100 % de rayonnement relatif. Les plus tolérants sont les sapins pectinés du Jura, les moins tolérants sont les mélèzes d'Europe (Alpes du Nord). On remarquera qu'à cet âge, les sujets, bien que dans leur période de phototropisme juvénile, manifestent déjà des réactions assez nettes aux variations de l'intensité du rayonnement naturel. A 2 ans, les différences entre les espèces sont encore plus marquées (figure 39), mais, sous 6% de rayonnement relatif, les mélèzes avaient disparu.
Après d'autres essais sur des espèces non indigènes (sapin de Vancouver, cèdre de l'Atlas, épicéa de sitka, douglas vert, séquoia géant, tsuga hétérophylle), effectués dans le Haut Jura pendant une année, et qui n'ont pu être suivis plus longtemps (ROUSSEL - 1955), une expérience plus importante et de longue durée a été organisée en Champagne humide. 12 caissettes de végétation comportant chacune 12 cases ont été installées dans une grande clairière et sous 5 peuplements de densité différente (2 caissettes par station), remplies d'un sol identique. 100 graines de chacune des 12 espèces résineuses ont été semées, dans chaque case, au printemps 1962 ; voici leur liste, avec, en référence, le numéro de classement qui sera reporté dans le tableau général ci-dessous : (1) sapin pectiné (Jura) - (2) sapin pectiné (Aude) - (3) sapin de Vancouver (reboisements français) - (4) sapin de Nordmann (reboisements français) - (5) mélèze du Japon (Japon) - (6) épicéa commun (Jura) - (7) épicéa commun (Alpes du Sud) - (8) épicéa omorica (reboisements français) - (9) épicéa de sitka (U.S.A.) - (10) pin laricio de Corse (Corse) - (11) pin Weymouth (Vosges) - (12) douglas (Vancouver). Les stations allaient du plein découvert (Rr = 100 %) à la grande trouée (Rr = 70 % en été), à la petite trouée (Rr = 35 % en été), à la " coupe d'abri " (16 % de Rr en été), et aux taillis denses, très faiblement éclaircis (4 % de Rr en été), ou non éclaircis (3 % de Rr en été) (Fig. 16). La germination des graines, quelle que soit leur espèce, a été en général satisfaisante pour l'ensemble des stations (à l'exception des sapins, dont le taux de germination est, très constamment, assez faible), mais surtout aux environs d'un Rr de 35 %,

(papier millimétré).
De gauche à droite : sapin pectiné, épicéa
commun, pin à crochet, pin noir, pin sylvestre,
mélèze d'Europe.
De haut en bas : rayonnements relatifs de 6%, 12%, 37%, 100% (ROUSSEL
1953).
Les premières mensurations ont été effectuées au bout de 3 ans (ROUSSEL - 1965) et sont reproduites, pour certaines espèces caractéristiques, à la figure 35. On notera d'abord que sous des Rr de 3 et 4 % toutes les espèces ont disparu rapidement, malgré une bonne germination, après une dizaine d'essais successifs (7 portant sur les 12 espèces et 3 portant uniquement sur les sapins). Il s'agissait probablement d'une banale " fonte des semis ". A 4 ans, les différences de réactions entre les variétés et les espèces se sont accentuées (Fig. 36, 37, 38). On remarquera surtout la différence de développement entre les sujets installés sous des Rr de 16 % et 35 % - alors qu'entre 35% et 70% ces différences s'atténuent.
))
FIG. 35 - Corrélations observées
entre le rayonnement relatif des sous-bois feuillus, en été, et
la hauteur de 6 résineux âgés de 3 ans, qui s'y développent,
en cases de végétation.
NOTA: La disparition du sapin pectiné du Jura, à partir de
35%. de Rr, est due à un été exceptionnellement chaud et
sec. (ROUSSEL 1965).
En ce qui concerne le nombre des espèces se développant simultanément d'une manière convenable, le Rr de 35 % a semblé le plus favorable ; au-dessus, une sélection s'opère entre les sujets, certains continuant à bénéficier d'un rayonnement plus intense. Il faut noter, également, que dans la grande trouée (abritée des vents du Nord et de l'Est, et soumise à une insolation intense), on a observé, en 1964 surtout, un " coup de chaleur " qui s'est manifesté d'une façon bien moins marquée dans les stations de plein découvert (réduction des phénomènes de transpiration ?).
))
FIG. 36 - Etat de 12 résineux (voir
le texte) sous une coupe d'abri un peu sombre (Rr = 16%
en été), à l'âge de 4 ans. Les chênes pédonculés,
naturels, ont le même âge (quadrillage
0, 15 x 1, 15 m) (ROUSSEL 1966).
))
FIG. 37 - Etat de 12 résineux (voir
le texte) dans une petite trouée (Rr = 35 % en été),
à l'âge
de 4 ans. Les chênes pédonculés, naturels, ont le même
âge. (quadrillage 0,15xO,15 m).
(ROUSSEL 1966).
))
FIG. 38 - Etat de 12 résineux (voir
le texte) dans une grande trouée (Rr = 70%. en été)
à l'âge de 4 ans (quadrillage 0,15xO,15 m) (ROUSSEL 1966).
Cette expérience a été suivie en forêt jusqu'à l'âge de 8 ans, et au cours des 7e et 8e années, la mesure d'un certain nombre de facteurs physiques a été effectuée, en été et en hiver. Voici le type de corrélation que l'on peut observer, à l'âge de 8 ans, entre les divers sujets, le rayonnement relatif, l'humidité du sol et le pH en été :
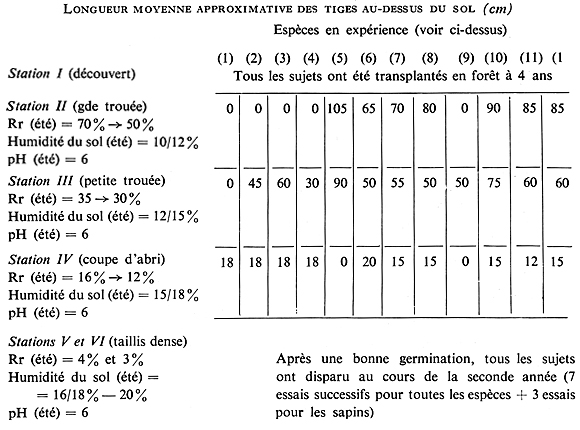
NOTA. Le coefficient de corrélation entre la hauteur de tous les résineux en expérience (concordant, du reste, avec leur développement général, visible à 4 ans sur les figures 36, 37 et 38), et le rayonnement relatif, en été et en hiver, est de l'ordre de + 1 (liaison positive absolue). Le coefficient de corrélation du même élément avec le pourcentage d'humidité du sol, en été et en hiver, est de l'ordre de -1 (liaison négative absolue) et avec le pH, de 0 (aucune liaison significative). Il en résulte que le développement des résineux est lié, positivement, d'une façon hautement significative (s°°°) à l'intensité du rayonnement naturel reçu au sol, sous réserve de l'action éliminatoire absolue exercée par cette intensité quand elle atteint une valeur trop élevée (cas des sapins - étés très chauds et très secs).
Ces résultats sont à rapprocher de ceux obtenus au Canada,
dans la région de Chalk River, par LOGAN (1959), sur le pin Weymouth,
développé en cases de végétation à sol identique.
La variation simultanée des divers facteurs physiques a été
reproduite à la figure
27. Les cases, au nombre de 40 (4 répétitions pour 5 degrés
de rayonnement relatif, avec des sujets âgés de 4 ans, élagués
et non élagués) ont été suivies pendant 4 ans. Voici,
pour les seuls sujets non élagués (plus proches de l'état
naturel), quelques-uns des résultats obtenus :
|
Rayonnement relatif (Rr)
|
Développement des sujets
après 4 années d'observations
|
Poids sec produit en 4 ans
|
|||
|
Hauteur de la tige (cm)
|
Diamètre pousse (mm)
|
Diamètre collet (mm)
|
cimes (gr)
|
racines (gr)
|
|
|
100%
|
102
|
5
|
25
|
277
|
86
|
|
55%
|
93
|
4
|
19
|
132
|
51
|
|
27%
|
75
|
3
|
13
|
48
|
20
|
|
19%
|
78
|
2
|
13
|
69
|
17
|
|
14%
|
60
|
2
|
10
|
32
|
13
|
Les tests statistiques calculés par
LOGAN, entre diverses caractéristiques des sujets, et les facteurs physiques
de chaque station montrent que c'est le rayonnement relatif qui, de beaucoup,
s'avère comme étant le plus important pour déterminer la
croissance du pin Weymouth (en poids, jusqu'à 100 % ; en hauteur
jusqu'à environ 50 %). La relation positive avec les autres facteurs
(hydriques en particulier) est bien moins marquée, parfois même
absolument nulle.
FAIRBAIRN (1966-1967) a étudié, en cases de végétation à sol identique, sous des Rr de 100 %, 50 %, 25 %, 12,5 % et 6,25 % les réactions de certains résineux utilisés dans les reboisements en Grande-Bretagne (Ecosse). Il s'agissait de : (1) épicéa commun, (2) épicéa de sitka, (3) sapin pectiné, (4) sapin de Vancouver, (5) douglas, (6) tsuga. À 1 an, il observe des types de réactions très voisins de ceux reproduits à la figure 34. À 2 ans, il obtient les résultats suivants :
|
Rr
|
Longueur de la tige en cm
|
Longueur de la racine en cm
|
||||||||||||
|
(1)
|
(2)
|
(3)
|
(4)
|
(5)
|
(6)
|
(1)
|
(2)
|
(3)
|
(4)
|
(5)
|
(6)
|
|||
|
100%
|
14
|
23
|
8
|
18
|
29
|
20
|
21
|
23
|
20
|
20
|
23
|
22
|
||
|
50%
|
15
|
18
|
8
|
21
|
31
|
22
|
19
|
17
|
16
|
19
|
19
|
18
|
||
|
25%
|
15
|
16
|
9
|
18
|
26
|
20
|
17
|
16
|
17
|
19
|
18
|
17
|
||
|
12,5%
|
11
|
12
|
8
|
14
|
16
|
13
|
13
|
12
|
16
|
16
|
14
|
12
|
||
|
6,25%
|
9
|
9
|
7
|
9
|
15
|
10
|
10
|
10
|
14
|
12
|
13
|
10
|
||
Jusqu'à 25 % de Rr, la relation
entre l'accroissement en hauteur de la tige (ainsi que le développement
général) et l'intensité du rayonnement naturel est tout
à fait frappante. Au-dessus, l'accroissement en hauteur est plus irrégulier,
mais le développement d'ensemble reste lié à cette intensité.
En ce qui concerne le développement
des racines, le même type de réaction est observé, et plus
régulier, en général.
En ce qui concerne le poids sec total (tige et racine), on trouve une liaison absolument constante avec l'intensité des radiations reçues par les sujets - sauf une légère inversion dans le cas du sapin pectiné (entre 25 et 50 %) :
|
Rr
|
Poids sec total (tige et racine)
en grammes
|
||||||
|
(1)
|
(2)
|
(3)
|
(4)
|
(5)
|
(6)
|
||
|
100%
|
4,8
|
14,1
|
4,5
|
8,3
|
23,5
|
11,1
|
|
|
50%
|
3,7
|
4,4
|
2,8
|
6,1
|
13,3
|
5,4
|
|
|
25%
|
2,6
|
2,9
|
3,1
|
4,4
|
5,4
|
3,0
|
|
|
12,5%
|
0,9
|
1,1
|
1,6
|
2,4
|
1,7
|
0,9
|
|
|
6,25%
|
0,6
|
0,5
|
1,0
|
1,2
|
1,2
|
0,5
|
|
En somme, on peut conclure de ces diverses observations, que le rayonnement naturel exerce une influence favorable à peu près certaine sur la croissance de tous les jeunes arbres résineux. Au début, la différence entre les sujets s'étant développés sous des Rr très différents est marquée, mais d'une façon non excessive, tout au moins pour certaines espèces. Puis, comme le très jeune arbre consacre une part importante des matières organiques qu'il synthétise à la construction de nouvelles aiguilles, on observe, d'année en année, une sorte d'effet cumulatif, et l'âge des sujets intervient, sous une forme exponentielle, dans le développement général, le poids frais et sec, la hauteur de la tige au-dessus du sol, la longueur des racines, etc ...

FIG. 39 - Sapins pectinés (ligne
supérieure) et Mélèzes d'Europe (ligne inférieure)
après
2 ans, développés sous 12% (à gauche), 37% (au
centre) et 100% (à droite) de rayonnement
relatif (Papier millimétrique) (ROUSSEL 1954).
Cependant, il ne faudrait pas en conclure, un peu hâtivement,
que l'on doive mettre tous les sujets, de toutes les espèces, dans
toutes les stations, immédiatement en pleine lumière. En effet,
un rayonnement naturel élevé, surtout parce qu'il s'accompagne
de chaleur, risque de dessécher à l'extrême les couches
superficielles du sol, de brûler même certaines jeunes tiges,
d'accentuer les phénomènes de la respiration, etc ... ; en somme
d'exercer une action limitante tout aussi importante que celle qui résulte
d'une nette déficience en radiations. On tirera de cette remarque certaines
conclusions pratiques, à la page
127 ci-dessous. Enfin, on remarquera qu'il n'est pas absolument certain
que les sujets issus de graines se développent de la même façon,
dans la station où ils ont été semés, que des
sujets âgés de quelques années introduits pour des plantations,
dans cette même station.
![]()