Article publié en 1969 dans la Revue forestière française, n° 1, pp. 27-32 - (télécharger la version .pdf - 644 k)
Le très jeune épicéa et la lumière
par Louis
ROUSSEL
(Les chiffres placés
dans le texte, entre parenthèses, renvoient à la bibliographie.)
Pendant leur jeunesse, en
général au cours des 1ère et 2ème
années de leur croissance, de nombreux résineux, communs dans
notre pays, sont freinés dans leur élongation par la lumière.
Ils réagissent par une courbure phototropique, quand l'éclairement
est unilatéral (voir ci-dessous), ou par un ralentissement général
de leur croissance, quand cet éclairement est bien également
réparti. Par voie de conséquences, l'adjonction d'un abri circulaire
latéral accélère leur élongation. Ces propriétés
disparaissent, la plupart du temps au cours de la 3ème année,
dans les conditions naturelles, et, à ce moment, le développement
de ces résineux est assez intimement lié à leur nutrition
carbonée, donc à l'intensité, constamment changeante
dans le milieu forestier, de la lumière qui atteint leurs aiguilles
(1).
On a pensé qu'il était
possible de différencier, très précocement, les races
de pin sylvestre, en étudiant, minutieusement, leur courbure phototropique,
dans une lumière artificielle unilatérale, en général
de faible intensité (2). Si certaines races, ou variétés,
sont plus sensibles que d'autres à la lumière latérale,
elles devraient logiquement s'allonger davantage en abri circulaire.
Note. - Les causes
véritables du phototropisme restent encore assez mystérieuses,
bien que, depuis longtemps, diverses explications aient été
proposées par les Botanistes RAY, au 17ème
siècle, HALES au 18ème , qui pensaient surtout à
une différence de température entre les côtés,
éclairés et ombragés. MIRBEL, dont les idées furent
souvent admises par les Forestiers français au début du 19ème
siècle, estimait que l'obscurité, entretenant la souplesse des
parties végétales, favorisait leur allongement, cependant que
la lumière, en aidant à leur nutrition, les consolidait et arrêtait
leur croissance. La taille réduite des végétaux de haute
altitude était expliquée de cette façon.
Préparée par
les expériences de C. et F. DARWIN, et de BOYSEN JENSEN, la théorie
proposée par WENT (1928) ouvrit des horizons nouveaux, et ce dans de
nombreux domaines de la physiologie végétale. La croissance
des plantes étant sous la dépendance d'hormones qui agissent
à dose extrêmement faible, l'une d'elles, l'auxine (on sait maintenant
qu'il s'agit de l'hétéro-auxine) est déplacée
par la lumière vers le côté ombragé. D'où
une croissance plus active de cette partie du végétal, entraînant
la courbure phototropique. Mais, même en ce qui concerne le coléoptile
d'avoine (sorte de gaine très fugace enveloppant la première
feuille au moment de la germination, et qui a été, pour des
raisons pratiques, spécialement étudiée), d'autres expérimentateurs
ont mis en évidence une inactivation, ou une destruction de l'hétéro-auxine
(directement, ou par l'intermédiaire de substances photosensibilisatrices),
On a pensé aussi à une action nuisible de la lumière
sur des enzymes, ou sur des co-facteurs d'auxines et même à un
effet ralentisseur direct sur l'activité des cellules des méristèmes.
La théorie complète du phototropisme est donc encore en voie
d'élaboration (bien que la sensibilité de l'hétéro-auxine
à la lumière paraisse définitivement admise). L'auteur
de ce mémoire estimant que la courbure des jeunes résineux vers
la lumière, et leur allongement en abri circulaire latéral,
ne sont que les deux aspects d'un même phénomène (puisqu'ils
apparaissent, et disparaissent en même temps, et qu'ils semblent, par
ailleurs, logiquement liés), Il lui est donc impossible de formuler
une explication physiologique précise, compte tenu des incertitudes
qui viennent d'être exposées.
Le présent travail
a eu pour but d'étudier, non la courbure phototropique, mais l'allongement
produit par un abri circulaire standard (3,5 cm de hauteur), en lumière
naturelle, chez deux variétés d'Epicéa commun (Picea
excelsa Link. = Picea Abies Karst.). Les graines, d'origine contrôlée,
ont été fournies par M. BOUVAREL, Directeur de Recherches au
C.N.R.F. Voici les caractéristiques de ces deux variétés
:
1er lot -
No 63103 - Provenance Morzine (Alpes du Nord).
2ème lot
- No 58 305 - Provenance St-Laurent II (Jura).
Les essais ont eu lieu dans
une petite serre bien éclairée naturellement (l'intensité
lumineuse y atteignait, par moments, de 30 à 50.000 lux sur une surface
horizontale, à partir du milieu du mois d'avril 1968) ; la température
y oscillait, à l'ombre, entre 15° et 20° C environ, et l'état
hygrométrique y est resté élevé (75 à 90
%). Le régime photopériodique naturel du lieu de l'expérience
présentait, pendant toute la durée de l'expérience, une
durée du jour légèrement supérieure à celle
des régions d'origine des deux variétés.
Deux séries de dispositifs ont été utilisées : pour les graines du premier lot (200 environ), 12 petits pots revêtus de couleur blanche, remplis de vermiculite, étaient répartis en 9 dispositifs, avec répétition des plus importants : (T) témoins ; (N) abri noir de hauteur standard ; (B) abri blanc de hauteur standard ; (2B) abri blanc de deux fois la hauteur standard. En outre, certains abris latéraux étaient, soit simplement ajourés (B/2), soit ajourés et munis de filtres colorés en bleu-outremer (B/0), vert (B/V), jaune (B/J) et rouge (B/R) (fig. 1).
 |
|
Fig.1 : Epicéa des Alpes. Ordre des dispositifs de gauche à droite : T, N, B/O, B/V, B/J, B/R, B/2, B, 2B. PHOTO ROUSSEL |
Pour les graines du deuxième lot (200 environ), 12 petits pots revêtus de couleur noire, remplis de vermiculite, étaient répartis en 8 dispositifs, avec répétition pour les plus importants - (T) témoins ; (N) abri noir de hauteur standard ; (2N) abri noir de deux fois la hauteur standard. En outre également, certains abris latéraux étaient, soit simplement ajourés (N/2), soit ajourés et munis de filtres colorés en bleu-outremer (N/O), vert (N/V), jaune (N/J) et rouge (N/R) (fig. 2).
 |
|
Fig. 2 : Epicéa du Jura.
Ordre des dispositifs de gauche à droite : T, N, N/O, N/V, N/J,
N/R, N/2, N, 2N. PHOTO ROUSSEL
|
La question de l'alimentation
en eau était spécialement traitée : chaque soir, on réglait
chaque dispositif de telle sorte que la moitié des vides du sol était
pleine d'eau, l'autre moitié restant pleine d'air. Une rotation biquotidienne
des dispositifs était pratiquée (l'éclairement, dans
la serre, était un peu déséquilibré), ainsi qu'une
lente interversion de ces dispositifs. Les sujets ont été extraits
à la fin du mois de juin 1968 et mesurés au mm près,
en distinguant la longueur des axes hypocotylés, et celle des radicelles.
Les chiffres obtenus ont été traités par la méthode
préconisée par STUDENT et FISHER, et décrite par J. PARDÉ
dans son récent Traité de Dendrométrie sous la dénomination
de " Comparaison des moyennes - cas des petits échantillons "
(3). Comme il restait, en général, au moins 10 sujets en bon
état de végétation, dans chaque dispositif, le calcul
du test " t " était tout à fait justifié.
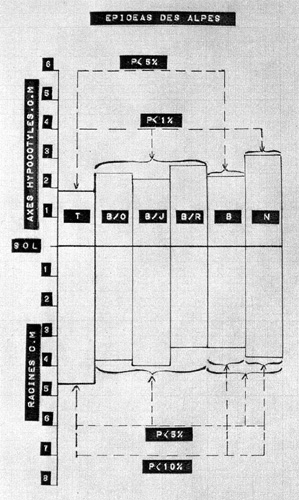
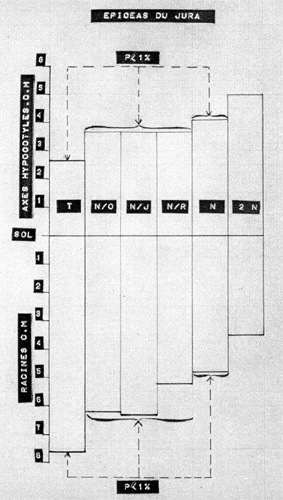
Les figures n° 3 et 4 exposent les résultats obtenus, pour chacune des variétés en expérimentation ; les indications P < 5 %, P < 1 %, indiquent que les différences observées entre les moyennes sont, selon l'usage, significatives, ou hautement significatives.
 |
 |
|
|
Fig. 3 Longueurs moyennes des axes
hypocotylés et des racines chez les Epicéas des Alpes,
suivant les dispositifs de protection. PHOTO
ROUSSEL
|
Fig. 4 Longueurs moyennes des axes
hypocotylés et des racines chez les Epicéas du Jura, suivant
les dispositifs de protection. PHOTO ROUSSEL
|
Pour commenter brièvement
ces deux figures, on soulignera :
1° L'efficacité,
très générale, de l'abri circulaire latéral sur
l'élongation de l'axe hypocotylé.
L'abri noir est, pour la variété des Alpes du Nord, plus efficace
que l'abri blanc, en raison d'une légère diminution de l'opacité
du second par rapport au premier, et, également, d'une réflexion,
non négligeable, de sa paroi interne.
2° La différence
de longueur des axes hypocotylés entre les sujets des deux variétés,
sans aucun abri (T), ou avec abri noir (N), seuls cas exactement comparables.
Il est à noter que le principe de cette comparaison qui fait intervenir
le tout premier stade de la germination
(phase hypocotylée), diffère de celui exposé récemment
par Ph. BARADAT (4) qui se réfère à la longueur de la
pousse primaire (phase épicotylée). Cet auteur relève
également des différences entre certaines variétés,
qui seraient dues, selon lui, à une variabilité, d'origine génétique,
dans l'activité photosynthétique des aiguilles.
3° Le peu d'importance
que revêt la couleur de la lumière latérale, mais on peut,
dans ce cas , mettre en cause la façon dont étaient établis
les dispositifs expérimentaux.
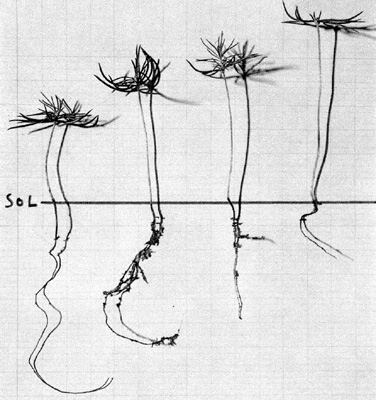
4° Le changement de la répartition des " facteurs trophiques de croissance " entre les axes hypocotylés et les racines, en fonction du dispositif de protection utilisé. Ce raccourcissement de la longueur des racines, en corrélation avec l'allongement des axes hypocotylés, est très net chez l'épicéa du Jura. Il est, du reste, assez logique. La graine d'épicéa est très petite, très légère (0,01 gramme en moyenne). Les matières plastiques existant en quantité limitée, au moment de la germination, leur " mobilisation, commandée par la plus ou moins grande abondance des substances de croissance de nature auxinique, à la construction de l'axe hypocotylé (tant que la concentration en hétéro-auxine reste supérieure à un certain seuil), entraîne inévitablement une modification du reliquat disponible pour la formation de la radicelle (laquelle, au surplus, ne s'accroît qu'avec une concentration en hétéro-auxine inférieure à ce seuil). Or, et bien qu'en cette matière complexe, on doive se montrer très circonspect (on l'a bien vu plus haut), on ne peut manquer de remarquer que c'est sous un éclairement latéral élevé (réduisant peut-être rapidement les substances auxiniques actives), que l'on enregistre une croissance limitée de l'axe hypocotylé, et un développement important de la radicelle. Au contraire, sous un éclairement latéral réduit (permettant le maintien d'une concentration auxinique assez élevée), on observe la formation d'un axe hypocotylé assez long, utilisant une quantité relativement importante de matières plastiques, et n'en laissant que très peu pour la construction de la radicelle, laquelle demeure courte (fig. 5).
 |
|
Fig.
5 Epicéas du jura de type moyen dans les dispositifs : T, N/O,
N, 2 N (de gauche à droite). |
Les réactions de l'Epicéa des Alpes du Nord sont de moindre amplitude que celles de l'Epicéa du Jura. Certes, on constate un allongement moyen général des axes hypocotylés en abri circulaire, mais le raccourcissement des radicelles est nettement plus irrégulier, Certains sujets, même ombragés, conservent des racines assez longues (la variance est élevée, ce qui explique la moins bonne signification statistique de la différence entre les moyennes). Cette observation justifierait, à elle seule, s'il en était besoin, les très intéressants travaux poursuivis depuis de nombreuses années sous la direction de P. BOUVAREL (5), au sujet de l'importance des provenances, pour l'utilisation de cette remarquable espèce ligneuse.
 |
Fig. 6 Epicéas communs (groupes de gauche) et Sapins pectinés (groupes de droite) du Haut Doubs, à la fin de leur première année de croissance, dans leur milieu forestier d'origine. Rangée supérieure : sous un perchis très dense (1,22 % de Rr). Rangée moyenne : sous futaie dense (3,28 % de Rr). Rangée inférieure
: dans une grande clairière (98 % environ de Rr). |
À côté
de leur signification physiologique, ces observations ont de nombreuses conséquences
culturales. En effet, l'architecture générale de la plantule
(surtout pour la variété du Jura), déterminée
lors de la germination par les conditions de l'éclairement, se retrouve,
dans le milieu naturel, et se maintient,
approximativement, pendant toute la première année de sa croissance.
Le jeune sujet, développé dans une trouée bien isolée,
conserve la longueur réduite de son axe hypocotylé, à
laquelle s'ajoute celle de sa courte pousse primaire, lorsque les cotylédons
et les premières aiguilles ont commencé leur activité
photosynthétique. Mais la radicelle continue à s'allonger, et
parfois à se ramifier, jusqu'à la fin de l'automne. Les observations
inverses sont faites pour la plantule développée sous un peuplement
bas et fermé (fig. 6). On savait déjà, par ailleurs,
que la structure anatomique (coupes transversales) des axes et des cotylédons
varie considérablement suivant que le sujet s'est développé
à l'ombre dense, ou dans une très grande clairière (6)
: les formations libéro-ligneuses secondaires, les parois lignifiées
et la paroi épidermique externe étant beaucoup plus développées
dans ce dernier cas. Les conséquences pratiques sont importantes :
- Coupes d'ensemencement. Alors que l'on ne conteste pas l'intérêt
des coupes, secondaires ou définitives, pour assurer J'avenir d'une
régénération d'Epicéa, installée sous le
couvert, on a souvent douté du rôle de la " coupe d'ensemencement
" (préalable à la chute des graines et à leur germination).
La différence de structure entre la plantule de lumière, et
celle d'ombre, en particulier la longueur très dissemblable atteinte
par les radicelles dans les mois qui suivent la germination, montre, d'une
façon peut-être paradoxale, que c'est la première qui
résistera le mieux à un dessèchement superficiel du sol.
La seconde étant, sur ce point, bien plus sensible.
- Installation capricieuse
des régénérations.
Ce caractère, qui a fait parfois qualifier l'Epicéa " d'espèce
ondoyante et diverse ", et qui se manifeste, notamment, par les phénomènes
d'alternance fréquents dans le Jura, a reçu déjà
de nombreuses explications : concurrence radiculaire et toxicité de
la litière accumulant le manganèse assimilable (7), présence
d'inhibiteurs de germination de nature biologique
complexe (8), influence de la plus ou moins bonne mycorhization (9), abondance
des hautes herbes et modification des propriétés microbiologiques
des sols (10). Or, il semble qu'il convienne de faire entrer aussi
en ligne de compte le comportement du " principal intéressé ",
à savoir celui de l'Epicéa lui-même. Si l'on admet, avec
DUCHAUFOUR (7) que, dans certains cas, la couche éventuellement toxique
(ou desséchée) peut être de faible épaisseur (1
ou 2 cm), on comprend très bien que les plantules d'ombre, qui ont
des radicelles de cette longueur, dépériront dans ce milieu,
d'une façon irrémédiable. Par contre, les plantules de
lumière, dont les radicelles atteignent 7 à 8 cm en juin, davantage
en fin d'année, seront vite délivrées de cette néfaste
influence.
Comme certaines variétés
paraissent avoir des comportements différents, en particulier des radicelles
de longueur très irrégulière, même pour des sujets
d'ombre dense, on conçoit qu'un type de raisonnement, valable pour
l'Epicéa du Jura, n'est pas obligatoirement transposable à celui
des Vosges, ou de la Forêt Noire...
- Pépinières
volantes installées sous bois.
Des réserves doivent être faites sur la pratique, commune autrefois
dans certaines régions, des pépinières installées
sous des perchis (en général de sapin), dans une ombre assez
dense. L'aspect des plantules est en général excellent, la première
année, mais leur structure, comportant de très courtes radicelles,
les rend très sensibles aux variations de l'humidité du sol.
- Généralisation
des conclusions. - Tout ce qui vient d'être dit pour l'Epicéa
du Jura s'applique aux espèces résineuses, à germination
printanière et épigée. Les Pins (sylvestre de Champagne,
laricio noir d'Autriche et maritime des Landes), se comportent - et d'une
manière parfois encore plus marquée - de la même façon,
selon qu'ils se développent dans une trouée, ou sous un couvert
dense. Ces commentaires ont donc une portée sylvicole assez générale,
même dans les régions méditerranéennes, à
climat lumineux et sec (11).
BIBLIOGRAPHIE
(1) ROUSSEL L. (1966). - Le phototropisme juvénile des résineux. R.F.F. n°11.
(2) SCHRÖCK
0. (1958). - Die Untersuchung der phototropischen Reaktion als Auslesemethode
bei Kiefersämlingen auf Gradschäftigkeit. Der Zucher, Band
28. Heft 7.
(3) PARDÉ J. (l96l). - Dendrométrie (page 40). École Nationale des Eaux et Forêts, Nancy.
(4) BARADAT Ph. (l967). - Élongation de la pousse primaire chez Picea abies en relation avec la date d'aoûtement et la teneur en chlorophylle des feuilles. Groupe d'Etude des Problèmes de Physiologie de l'Arbre, séance du 17 novembre 1967.
(5) BOUVAREL P. (1955). - La sélection individuelle des arbres forestiers. R.F.F. n°11.
(6) TRONCHET A. et ROUSSEL L. (1954). - Observations sur la structure anatomique de plantules d'Epicéa exposées à des conditions différentes d'éclairement. Ann. Scient. Univ. Besançon, 2ème série, Bot. 1.
(7) DUCHAUFOUR Ph. et ROUSSEAU L.Z. (1959). - Les phénomènes d'intoxication des plantules de résineux par le manganèse dans les humus forestiers. R.F.F. n° 12.
(8) SCHAEFFER R. et MOREAU R. (1958-1959). - L'alternance des essences. B.S.F.F.C., tome XXIX, n°1, 2 et 5.
(9) BOULLARD B. (1960-1962), - Importance des mycorhizes pour la régénération de l'Epicéa (Picea excelsa). B.S.F.F.C., tome XXX, n°1 ; tome XXXI, n°1 et 5 ; tome XXXII, n°1.
(10) MOREAU R. et POLY J. (1968). - La régénération de l'Epicéa dans les forêts de haute altitude. B.S.F.F.C., tome XXXIV, n°1 et 2.
(11) MAGINI E. (1966). - Recenti progressi della fotologia forestale in Francia. L'italia forestale e montana. Anno XXI, n°1.