
Les auxines, agents essentiels de la croissance des végétaux
par Louis Roussel
Article publié en 1974 dans la revue Bois et Forêts des Tropiques, n° 158, p 51 - (version .pdf - 513 k)
| SYLVICULTURE ET PHYSIOLOGIE VÉGÉTALE | |
|
Les auxines, agents essentiels de la croissance des végétaux par Louis Roussel
|
|
| Photo Brunck. - Teck. Après éclaircie, développement de gourmands sur un arbre à l'intérieur d'une parcelle. Bamoro, Côte d'Ivoire. |
SUMMARY
THE AUXINS, ESSENTIAL AGENTS IN VEGETABLE GROWTH
The auxins, particularly A. I. A. (acetic indol ß acid) determine the
growth of vegetable structures in accordance with certain laws; it is more particularly
phototropism which has allowed of the discovery and isolation of A. I. A.
The author studies the constitution of A. I. A. and its molecular action, the formation and destruction of A. I. A. and its effects on vegetation, the part played by A. 1. A. in geotropism and the other types of morphoses in which A. 1. A. and might play a part.
Dans une chronique précédente (1), on a rappelé la distinction,
très importante, entre les matières " plastiques ",
éléments avec lesquels les végétaux se construisent,
et les matières "oligodynamiques ", dont le rôle paraît
être de mettre en œuvre ces éléments, d'une façon
ordonnée, afin d'aboutir à des structures harmonieuses et équilibrées.
Parmi ces matières oligodynamiques, une place à part doit être
faite aux "auxines ", et tout spécialement à l'AIA (acide
ß indole acétique) dont on retrouve très fréquemment
la présence en physiologie végétale. Il est à noter
que, primitivement, l'AIA était appelé " hétéro-auxine
" ; il fut identifié ensuite comme étant l'acide ß
indole acétique, et, assez récemment, divers physiologistes ont
proposé de l'appeler simplement " auxine ". Pour éviter
toute confusion, car de nombreuses substances naturelles ou artificielles ont
des propriétés très voisines, on le désignera, dans
l'exposé qui va suivre, sous le sigle d'AIA (IAA dans les pays de langue
anglaise).
1. - DÉCOUVERTE DE L'EXISTENCE DES AUXINES
Le phénomène qui a conduit d'abord à la notion d'hormone
végétale, puis à l'isolement de l'AIA est, incontestablement,
le phototropisme (autrefois appelé héliotropisme) ; il s'agit
d'un processus assez général de la croissance des végétaux,
qui consiste en l'orientation des tiges, des feuilles, et parfois d'autres organes,
vers la lumière. En quelques lignes, on rappellera que, dès l'année
1880, C. et F. DARWIN, opérant sur un coléoptile de graminée
(sorte de gaine très fugace protégeant la feuille cotylédonaire
lors de la germination), avaient montré que c'était l'extrémité
de cet organe qui percevait le stimulus lumineux (unilatéral, dans le
phototropisme), mais que c'était la croissance (dissymétrique)
d'une région un peu inférieure qui entraînait, en définitive,
la courbure du coléoptile vers la lumière. D'où l'idée
d'un " messager chimique ", responsable de cette action à distance,
d'une façon un peu analogue à ce qui était constaté,
vers cette époque, en physiologie animale.
Par la suite, divers chercheurs (BOYSEN-JENSEN, PAAL, SÖDING, notamment)
préparèrent la voie qui conduisit à la première
théorie cohérente : celle qui porte, en général,
les noms de CHOLODNY-WENT, lesquels, autour des années 1926-1928, en
jetèrent les bases. C'est la répartition, inégale sur les
faces, éclairée et ombragée, d'une " substance
de croissance", sensible à l'action de la lumière, qui provoque
une élongation différente des cellules de ces deux faces, d'où
résulte la courbure observée du coléoptile.
Une remarque importante doit être faite à ce sujet : c'est sur
un organe mineur et transitoire qu'ont été effectuées la
majorité des premières expériences, et c'est au phototropisme
du coléoptile d'avoine que s'appliquent, évidemment, les théories
explicatives. Or il est certain que si le phototropisme se manifeste chez de
nombreux végétaux, et que si l'on y décèle facilement
l'intervention de l'AIA, les modalités de ce phénomène
peuvent varier sensiblement d'une espèce à l'autre ; ne serait-ce
que si l'on considère que certaines graines ont une germination hypogée,
et d'autres une germination épigée. Sur le coléoptile d'avoine,
le phototropisme est observé pendant quelques dizaines d'heures, au maximum
; chez les végétaux ligneux, il peut se manifester pendant de
très nombreuses décennies. Il faut donc faire preuve de beaucoup
de prudence en examinant les résultats des expériences effectuées,
et les théories élaborées, quand il s'agit de les étendre
à l'ensemble du règne végétal. Cette idée
sera reprise un peu plus loin.
Une autre remarque est à faire au sujet des expériences de SÖDING,
cité plus haut : dès l'année 1923, il avait montré
que, toujours chez le coléoptile de graminée, l'allongement de
cet organe à l'obscurité avait la même origine que la courbure
: l'action d'une substance de croissance, sensible à l'action de la lumière.
Les théories ultérieures ont surtout retenu l'effet "
courbure ", et très peu l'effet " allongement ".
Actuellement, quand l'on constate ce second effet, on pense la plupart du temps
à l'action obligatoire du phytochrome (1).
2.-CONSTITUTION CHIMIQUE DE L'AIA, MODE D'ACTION DES AUXINES AU NIVEAU MOLÉCULAIRE.
WENT avait, pour détecter la présence des substances de croissance, d'abord indéterminées, mis au point le " test Avena ". Un coléoptile d'avoine, d'une variété fixée, décapité avant d'avoir atteint sa taille définitive, recevait sur une partie de son extrémité un petit bloc de gélose renfermant la substance à étudier. Si cette substance était active, et diffusait de haut en bas, la région située sous le bloc de gélose s'allongeait plus que l'autre, et il en résultait une courbure du coléoptile. Plus cette courbure était marquée, plus la substance était active. Divers extraits biologiques étaient capables de produire ce phénomène, en particulier l'urine humaine. KÖGL et HAAGEN-SMIT, en 1934, isolèrent dans ce produit plusieurs substances qui reçurent le nom général d'auxines. En fait, seule l'une d'elles se révéla stable et active: l'AIA dont la formule développée est donnée ci-dessous, et qui fut, on l'a dit plus haut, appelée "hétéro-auxine " à l'époque, car elle différait de deux autres substances que l'on avait, d'abord, considérées, sans doute à tort, comme les véritables auxines a et b.
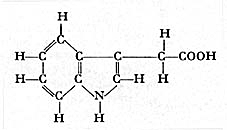
Acide ß indole acétique ou AIA.
Cette formule est très voisine de celle d'un acide aminé, le
tryptophane, abondant chez les végétaux. Il suffit, en effet,
de remplacer le groupement COOH de la formule ci-dessus, par le groupement suivant :
![]() ,
pour passer de l'AIA au tryptophane.
,
pour passer de l'AIA au tryptophane.
D'où l'idée, assez généralement admise, que le
premier est dérivé du second. Et comme la synthèse du tryptophane
nécessite la présence, transitoire, de zinc, en quantité
infinitésimale du reste, on comprend mieux pourquoi certains sols, fortement
carencés en zinc, sont très défavorables à la croissance
des végétaux - et pourquoi un apport, même minime - de cet
élément, peut dans ce cas avoir d'heureux effets sur la végétation.
L'exemple bien connu des forestiers australiens est tout à fait démonstratif
à cet égard.
De nombreuses autres substances, de synthèse parfois, jouissent des
mêmes propriétés que l'AIA quand on les soumet au "
test Avena ". Il semble que leurs caractéristiques chimiques
communes soient :
- de posséder un noyau cyclique comprenant au moins une double liaison.
Dans ce noyau, certains atomes de chlore peuvent très bien, du reste,
prendre la place d'atomes d'hydrogène ;
- d'avoir une chaîne latérale acide d'une certaine longueur (ni
trop courte, ni trop longue)
- de revêtir une certaine configuration spatiale conformément
aux principes de la stéréochimie.
Certains auteurs (WAIN) estiment actuellement que, pour qu'une substance ait
une activité auxinique, il faut qu'elle puisse " s'accoler "
à un accepteur, placé sans doute dans le cytoplasme de la cellule,
par 3 points : le noyau cyclique, le carboxyle, et un atome d'hydrogène
de la chaîne latérale. D'autres physiologistes (MUIR et coll.)
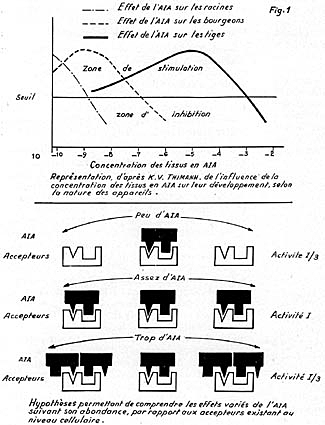
pensent que 2 points suffisent. La figure 1, représente approximativement
le mode d'action de l'AIA dans ce second cas. Ces concepts sur le rôle
de la configuration spatiale des molécules physiologiquement actives
(hormones, à action éloignée, ou enzymes, à action
rapprochée), sont actuellement admis dans beaucoup de phénomènes
biologiques, et ils en expliquent la très grande efficacité. On
trouvera de nombreux détails sur les théories relatives au mode
d'action des auxines dans un petit ouvrage très documenté de RICARD
(1960) (2).
De toutes façons, on estime que l'AIA, fixé sur son accepteur, intervient dans l'activité des cellules végétales en augmentant la plasticité des membranes, en accélérant l'absorption de l'eau par ces membranes, en stimulant le métabolisme général, et en particulier la respiration, etc...
3. - FORMATION, DESTRUCTION ET EFFETS LOCALISÉS DES AUXINES SUR LA
VÉGÉTATION.
Chez le coléoptile d'avoine, fragment isolé d'un végétal,
l'AIA est élaboré à l'apex et diffuse, de haut en bas,
dans l'ensemble de cet organe, à une vitesse de l'ordre de 1 à
2 cm par heure. Il semble qu'il en soit de même pour certaines plantules
entières, chez lesquelles l'AIA est élaboré dans les tissus
actifs de l'extrémité apicale, pour diffuser ensuite, de haut
en bas, jusqu'aux racines. Mais les feuilles, jeunes surtout, et plus tard le
cambium (assise de cellules très actives situées entre le bois
et le liber) sont également le siège de la production de l'AIA
(3). Dans d'autres cas, l'AIA est élaboré dans les cotylédons,
et, dans le cas de germination épigée, doit alors circuler de
bas en haut. En fait, DAVID, dans son petit ouvrage sur les hormones végétales
cité précédemment (1), relatant en particulier les travaux
de BOUILLENNE et de PALM-QUIST, estime que, dans une plante entière,
la circulation de l'AIA peut se faire dans tous les sens.
Cependant, il est très probable que des renseignements extrêmement
précieux seront fournis sur ce problème, grâce à
l'emploi de l'AIA " marqué " au carbone 14, appliqué
sur des feuilles, des bourgeons, etc.... ou injecté dans divers organes
des végétaux. On peut ainsi suivre les déplacements de
cette hormone, grâce à des compteurs du type GEIGER-MULLER, ou
bien par autoradiographie. En France, BONNEMAIN et ses collaborateurs ont commencé
très récemment des recherches dans cette direction, et les résultats
obtenus sont des plus prometteurs.
Comme l'AIA semble " s'user " très peu lors de son intervention
dans les phénomènes de croissance, et qu'il est produit en permanence,
il devrait s'accumuler dans les tissus. En fait, on ne le rencontre, en général,
que dans une proportion infime (en poids, de l'ordre du millionième,
ou du milliardième) ; ceci est dû à des phénomènes
antagonistes, également permanents, que l'on peut classer en deux catégories:
- la fixation des molécules d'AIA sur d'autres molécules organiques formant un ensemble provisoirement inactif, une sorte de mise en réserve (SIEGEL et GALSTON, 1953) ;
- la destruction des molécules d'AIA par oxydation, sous l'influence d'une enzyme : l'auxine-oxydase (TANG et BONNER, 1947). Cette destruction a été obtenue expérimentalement " in vitro " par GALSTON et BAKER en 1949, à la lumière, en présence d'une substance photosensibilisante présente dans de nombreux tissus végétaux : la riboflavine. Ce type de réaction est appelé : effet photodynamique. Il se déroulerait schématiquement de la façon suivante :
AIA (actif) + oxygène + lumière + enzyme + riboflavine
= ß indole formaldéhyde (inactif) + eau + gaz carbonique
Mais, si la perte d'activité est certaine, les modalités de cette
réaction ont été discutées. On a pensé aussi,
plus récemment, à la transformation de l'AIA, sous l'action de
la lumière, en substance temporairement inhibitrice de la croissance
(STILL et Coll. 1963).
Selon certaines conceptions, l'excès d'AIA pourrait, à lui seul,
provoquer la formation de l'auxine-oxydase, réalisant ainsi un bon exemple
d'autocompensation, assez courante dans divers processus biologiques. On pense
ainsi, que la teneur en auxine-oxydase des tissus du collet des végétaux
est élevée, et ceci correspond au fait, assez bien établi,
que la concentration en AIA des cellules des racines est normalement beaucoup
plus faible que dans les tiges, en particulier dans les méristèmes
terminaux.
Outre la lumière, une température élevée (de l'ordre
de + 50 °C) peut provoquer, chez des graines, la destruction de
l'AIA.
Plusieurs substances chimiques ont une action inhibitrice sur la production et sur la circulation de l'AIA : l'acide transcinnamique, par exemple. D'autres ont une action stimulante, ou synergique, comme l'acide 2-3-5 tri-iodo-benzoïque.

Aussitôt après la coupe des taillis, les chênes
de taillis sous futaie ont un tronc lisse, sans aucune branche latérale
(à gauche). Cinq ans après la coupe (à droite) de nombreux
gourmands se sont développés et deviennent des branches véritables,
dépréciant le bois tout entier (à droite). L'effet est
ici très marqué car la parcelle a été livrée
au bétail et les rejets de taillis n'ont pas pu se développer.
Un caractère important du mode d'action des auxines, parfois controversé,
mais cependant admis par la majorité des physiologistes, est le suivant
: l'AIA n'intervient dans un processus physiologique donné qu'à
un degré de concentration, chaque fois bien fixé. La figure 1,
proposée par THIMANN en 1937, et reprise par cet auteur en 1955 (4),
puis en 1969, schématise assez ce genre d'influence. Elle présente,
comme ou le verra plus loin, un intérêt certain et permet de relier,
en un ensemble cohérent les résultats d'un certain nombre d'expériences
en laboratoire, ou d'observations dans le milieu forestier. Sans doute, certains
phénomènes échappent-ils, en apparence, au cadre de ce
schéma simple, et doit-on, pour les y faire rentrer, élaborer
des hypothèses complémentaires ? peut-être, la teneur en
AIA d'un organe ne constitue-t-elle que l'un des éléments qui
déterminent son développement ? Il n'en reste pas moins que le
schéma de THIMANN possède une valeur figurative qui s'étend
à de nombreux phénomènes de la vie des arbres.
Il est à noter que KOPFER (5), spécialiste de ces problèmes,
qui les expose d'une façon détaillée dans son récent
ouvrage, cite des expériences tendant à établir que l'apparition
et le développement des bourgeons adventifs (se développant sur
des fragments de tissus blessés), sont déterminés par des
concentrations encore inférieures à celles, déjà
très faibles, nécessaires à la croissance des racines (STOUGHTON).
D'une façon générale, les considérations sur le
mode de jonction des molécules d'AIA sur les accepteurs au niveau cellulaire,
et les variations possibles du nombre de ceux-ci suivant la nature des organes
en cause, permettent d'imaginer des dispositions qui stimulent, ou inhibent
la croissance, ainsi que l'on a tenté de le faire sur la figure 1.
Ces remarques étant faites, on admet que l'AIA peut avoir sur les végétaux,
seul, ou accompagné d'autres co-facteurs, et aux concentrations convenables,
les genres d'effets suivants :
- il agit sur l'auxèse, c'est-à-dire sur l'allongement des cellules
constituées des tiges, des branches, des pétioles des feuilles
et des racines. C'est cet effet qui a été le mieux établi
;
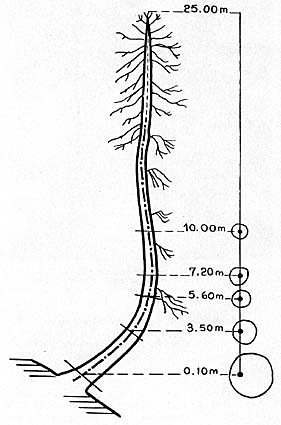
Vieux mélèze des Alpes dont la tige, d'abord
oblique, s'est redressée sous l'effet du géotropisme négatif.
Remarquez l'excentricité des cernes au cours du redressement géotropique.
D'après A. Engler, 1924.
- il agit sur la mérèse, c'est-à-dire sur la multiplication des cellules ; cet effet a surtout été mis en évidence sur la prolifération du cambium, la formation des cals sur les blessures. Mais il semble que cette action soit plus générale ;
- il peut provoquer la parthénocarpie, c'est-à-dire la formation
des fruits à partir de l'ovaire, sans fécondation par le pollen
;
- son insuffisance, à l'automne dans nos régions, entraîne
la formation d'assises cellulaires imperméables sur les pétioles
et les pédoncules, provoquant la chute des feuilles et des fruits;
- sa répartition dans l'ensemble des végétaux détermine,
sans doute en équilibre avec certains inhibiteurs, les " corrélations
de croissance ", spécialement étudiées par CHAMPAGNAT
(4), et il intervient aussi dans la "néoformation ~> des racines
et des bourgeons sur des organes tronçonnés ou blessés
;
- plusieurs auteurs estiment également que d'autres hormones végétales
récemment décrites (comme les gibberellines, agissant surtout
sur l'allongement des cellules, et les kinines, agissant plutôt sur leur
multiplication), n'agissent bien qu'en présence d'AIA, qui en augmente
sensiblement les effets ;
- enfin, on sait que les auxines de synthèse (surtout celles comportant
des atomes de chlore), à dose assez élevée, et parce qu'elles
ne sont pas détruites par l'auxine-oxydase, peuvent provoquer des malformations,
des nécroses... et que cette propriété est utilisée
culturalement, dans les désherbages par exemple. On pensera aux produits
désignés sous les noms de 2,4 D, de 2,4,5 T, de méthoxone,
etc...
Comme on le voit, les effets de l'AIA et des autres substances à propriétés
auxiniques, sont des plus nombreux et des plus variés.
4. - INTERVENTION, ORDINAIREMENT ADMISE, DE L'AIA DANS CERTAINS TROPISMES.
Les végétaux sont plus ou moins sensibles à divers facteurs
du milieu extérieur, et paraissent s'orienter pour les rechercher, ou
pour les fuir ; on citera parmi ces facteurs la pesanteur, la lumière
(ou certaines radiations invisibles), le frottement de contact avec un obstacle,
l'eau du sol, la présence de certaines substances chimiques, la différence
de potentiel électrique, etc... C'est ce que l'on appelle, en général,
des tropismes. Mais deux cas ont été spécialement étudiés
par les physiologistes : le géotropisme et le phototropisme. En ce qui
concerne les arbres forestiers, ENGLER (6) a effectué, il y a une cinquantaine
d'années, de nombreuses observations et expériences sur les conséquences
morphologiques et technologiques de ces tropismes.
Le géotropisme.
C'est l'orientation générale des racines dans le sens de l'attraction
de la pesanteur (géotropisme positif), et des tiges dans le sens contraire
(géotropisme négatif). Ce phénomène a depuis longtemps
été l'objet de l'attention des botanistes et sa réalité
est incontestable. Une preuve, indirecte, que le géotropisme est bien
dû à l'intervention de l'AIA est que si l'on bloque la circulation
de cette hormone par certaines substances inhibitrices mentionnées ci-dessus,
ce tropisme disparaît. On peut même, en dosant les effets, inverser
les réactions habituelles (racines se dirigeant vers le haut et tiges
se dirigeant vers le bas). RESPLANDY (1967) a effectué une intéressante
mise au point sur les recherches effectuées récemment dans cette
direction.
Quand on place un coléoptile d'avoine horizontalement, on trouve rapidement,
en utilisant le test de WENT, que le 1/3 environ de l'AIA total se trouve dans
la moitié supérieure, alors que les 2/3 sont rassemblés
dans la moitié inférieure. Il en résulte une croissance
différentielle des cellules du coléoptile, qui se redresse et
se dirige vers le haut. Pourquoi cette répartition différente
? la question semblait très claire, il y a quelques années, et
TRONCHET (7), en 1972, expose avec de nombreux détails les idées
d'HABERLANDT et de ses successeurs : des corpuscules extrêmement petits,
dénommés "statolithes " (en fait, on les assimile aux
grains d'amidon) se déplaçant lentement dans les cellules sous
l'action de la pesanteur, entraînent les molécules d'AIA, lesquelles
provoquent, par leur répartition différente, l'effet indiqué.
Cependant, ces idées ne sont pas universellement admises : on a objecté
que le phototropisme se manifeste, chez certains organes, en l'absence de grains
d'amidon et l'on a pensé à un déplacement des ions sous
l'influence d'une différence de potentiel électrique (effet géo-électrique
de BRAUNER).
Si l'on considère maintenant une racine placée horizontalement,
on observe la même différence de concentration des tissus en AIA
: nettement plus d'hormone dans la partie inférieure que dans la partie
supérieure. Pour expliquer que la racine se dirige vers le bas, on fait
intervenir la différence d'action de l'AIA, selon l'organe intéressé
et selon sa concentration (voir la fig. 1). Si, par exemple, la partie supérieure
de la racine a une concentration en hormone de 10-10 à 10-11
g/cm3 elle s'accroît. Mais si sa partie inférieure (qui est plus
riche en auxine) renferme une concentration de 10-8 à 10-9
g/cm3, sa croissance est stoppée. La racine se courbe donc vers le bas.
On ne doute plus guère, actuellement, que l'AIA ne soit, d'une façon
ou d'une autre, l'agent essentiel et obligatoire qui intervient dans le phénomène
du géotropisme. Il reste à trouver - si l'on n'accepte pas la
théorie des statolithes (malgré un nombre important d'observations
concordantes) - les raisons qui font que les molécules d'AIA sont, dans
un milieu riche en corpuscules de toutes sortes animés de mouvements
variés, attirés sélectivement par la pesanteur.
On doit remarquer - et ceci est valable pour tous les essais d'interprétation
qui vont être proposés plus loin - que la figure 1 (le schéma
de THIMANN) traduit bien plus un état de fait, un ensemble de constatations,
qu'elle ne constitue une explication fondamentale. Car, si l'on a observé
d'une façon fréquente que tel ou tel degré de concentration
en AIA agit de façon préférentielle sur le développement
de tel ou tel organe, les théories qui permettraient d'en comprendre
les raisons physiologiques profondes, ne recueillent pas une adhésion
unanime.
Le phototropisme.
Il est, on l'a dit, à l'origine des expériences qui ont abouti
à la découverte des auxines, à l'isolement de l'AIA, et,
ultérieurement, à la mise au point des hormones de synthèse.
On s'est beaucoup intéressé au coléoptile d'avoine, car
c'est un matériel vivant simple, peu coûteux, constant dans ses
réactions, et qui peut être rapidement mis en œuvre (en quelques
jours, on obtient des résultats utilisables). Mais ce phototropisme se
manifeste fréquemment dans le milieu forestier, non seulement sur des
semis âgés d'un ou deux ans, mais sur de jeunes arbres, et même
sur des tiges puissantes dont l'âge dépasse un siècle ou
deux, et qui ont, sur une partie de leur tronc, conservé l'orientation
spéciale qu'elles avaient prise dans leur jeunesse, sous l'influence
de conditions locales qui ont disparu par la suite.
Les anciens sylviculteurs l'avaient bien observé. BAUDRILLART, ce " compilateur consciencieux et infatigable ", rapporte dans son Traité Général des Eaux et Forêts publié en 1825, que l'orientation des troncs de chêne, vers la lumière, permettait d'obtenir des bois de marine de courbures variées et destinés à de multiples usages. Il semble même que cet effet ait été systématiquement utilisé aux XVIIe et XVIIIe siècle, par les forestiers royaux.
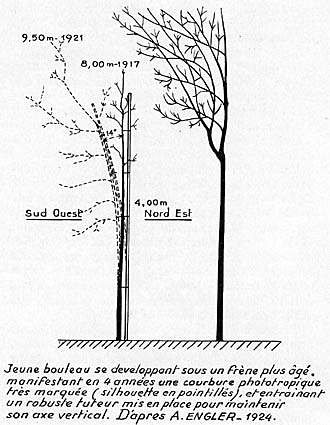
Pour en revenir au minuscule coléoptile d'avoine, des dosages d'AIA,
analogues à ceux effectués pour l'étude du géotropisme,
ont montré que cet organe, exposé à la lumière latérale,
renfermait environ 27 % d'hormone, du côté éclairé
et environ 57 %, du côté ombragé (par rapport à un
témoin, non éclairé, qui en renfermait par définition
100 %). Ces dosages mettaient en évidence une répartition différente
de l'AIA sur les côtés, éclairé et ombragé,
et également une disparition de 16 % environ de l'AIA primitif (ou supposé
tel, par rapport au témoin).
Pour résumer les interprétations auxquelles ont donné
lieu les expériences sur le phototropisme, on indiquera que la théorie
de CHOLODNY-WENT souligne, on l'a déjà dit, la répartition
différente de l'AIA - que GALSTON relève la disparition d'une
partie de cette hormone - et que BLAUUW et ses successeurs (GUTTENBERG en particulier)
insistent, au contraire, sur le ralentissement des facultés d'allongement
et de multiplication des cellules éclairées, ce qui renforcerait
considérablement les effets ci-dessus exposés. On trouvera un
exposé détaillé de ces différents points de vue
dans une étude de BRIGGS (6) parue en 1964.
Toutes ces théories doivent cependant tenir compte des faits suivants
: c'est la lumière bleue, présente dans la lumière naturelle
(de longueur d'onde allant de 340 à 500 nm, avec maximum d'efficacité
au voisinage de 440 nm) qui semble de beaucoup la plus efficace dans le phénomène
du phototropisme. Or, la courbe spectrale d'action de ces radiations correspond
assez bien à la courbe spectrale d'absorption par des pigments présents
chez le coléoptile d'avoine : des flavoprotéines tout spécialement.
D'où l'idée, précédemment exposée, que ces
substances pouvaient jouer le rôle de photosensibilisants. Au contraire,
l'effet photopériodique, on l'a dit précédemment (1), paraît
sous la dépendance de radiations de plus grandes longueurs d'onde (le
rouge clair et le rouge sombre).
Il est bien évident que si l'on ne peut expliquer parfaitement le mécanisme
du phototropisme, chez un organe d'aussi faible importance que le coléoptile
de graminée, qui se développe pendant une centaine d'heures, et
qui a été étudié depuis près d'un siècle,
on ne peut s'étonner des lacunes actuelles concernant les processus qui
se déroulent chez des arbres qui, pendant de très nombreuses saisons,
arrêtent, puis reprennent leur croissance, et sont soumis, sur l'ensemble
de leurs troncs, à une action plus ou moins déséquilibrée
de radiations variées, qui les pénètrent parfois profondément.
En effet, des mesures nombreuses effectuées depuis une quinzaine d'années
(papier photographique-microdensitomètre) ont montré que les écorces
d'arbres, même âgés, ne sont pas complètement opaques.
Chez des sujets de quelques années, on dépasse rarement des densités
optiques supérieures à 4 ou 5 (soit un facteur de transmission
optique de 1/10.000 à 1/100.000). Or, avec une densité optique
de 6, un centimètre carré de cambium reçoit, chaque seconde,
dans une tige bien éclairée, quelques 109 photons de
lumière visible, dont l'énergie individuelle varie de 1 à
3 eV. Sous ce même centimètre carré se trouvent, à
une concentration admissible de 10-5 à 10-10 g/cm3,
de 3. 1011 à 3.1016 molécules d'AIA. Qu'en
présence d'auxine-oxydase, et grâce à la flavoprotéine
photosensibilisante un certain nombre de puissances de 10 de ces molécules
soient oxydées, et perdent ainsi leur efficacité, n'est nullement
invraisemblable - ceci, en accord avec les processus admis dans de nombreux
domaines de la photophysiologie.
Ce raisonnement conduit à d'autres effets, encore mal connus dans tous
leurs détails, mais qui se rencontrent fréquemment dans le milieu
forestier. JACQUIOT (3) et CATINOT (9) en ont relevé récemment
un certain nombre dans les forêts des régions tempérées
ou tropicales ; la lecture de leurs études sera très utile pour
mieux comprendre les développements qui suivent.
5. - AUTRES TYPES DE MORPHOSES OÙ INTERVIENNENT, PROBABLEMENT, L'AIA
ET LA LUMIÈRE.
Cas des jeunes arbres dont les cimes sont maintenues en pleine lumière,
alors que leurs tiges sont ombragées plus ou moins fortement.
Si l'on dispose, en plein découvert, de jeunes chênes rouvres
et pédonculés, assez espacés pour que leurs tiges reçoivent
une lumière latérale suffisante, on obtient, au bout de quelques
années, des sujets courts, ramassés, et pourvus de nombreux rameaux
latéraux. Le fait d'ombrager latéralement les tiges seules
de certains d'entre eux, en laissant les feuillages bien éclairés,
provoque une sensible élongation. La forme des sujets abrités
latéralement se rapproche de celle des arbres de futaie - alors que les
sujets non abrités sont semblables à des arbres isolés,
de taillis sous futaie par exemple. La longueur individuelle des cellules qui
constituent les tiges reste la même, dans les deux cas : ce qui montre
que l'ombragement latéral a favorisé bien plus la multiplication
que l'allongement cellulaire. Mais la disposition horizontale des cellules est
un peu modifiée. Les cernes des sujets abrités (longs) sont, au
collet, plus étroits et plus riches en bois de printemps, et les rayons
ligneux y sont moins nombreux que chez les sujets non protégés.
Ce genre d'expérience est relativement facile à réaliser,
et la réaction d'allongement a été observée sur
des sujets âgés de 4 à 5 ans au maximum. DELVAUX, à
la station de recherches des Eaux et Forêts de Groenendaal, en Belgique,
a obtenu récemment un effet d'allongement net sur des boutures de peupliers
dont certaines étaient abritées par des écrans latéraux
de plastique noir.
Pour des sujets plus âgés, il faudrait pousser la protection latérale
sur une hauteur de plus en plus grande, ce qui n'est pas toujours facile pratiquement.
Mais il existe, dans le milieu naturel, des exemples de ce que peut donner la
protection latérale pendant une durée bien plus longue (cas de
certains arbres feuillus se développant dans des plantations résineuses
très denses, et qui croissent surtout en longueur, alors que leur diamètre
reste faible). Là encore, on constate que les dimensions longitudinales
des cellules des tiges restent les mêmes dans tous les cas (sujets "
longs " et sujets " courts ") (10).
Quand on procède de cette façon sur des sujets résineux très jeunes (pendant les mois qui suivent leur germination, alors qu'ils manifestent un phototropisme très net), on obtient, par l'ombragement latéral, des sujets à axes hypocotylés longs, et à radicelles courtes. Plus l'ombragement latéral s'atténue, plus les axes deviennent courts, alors que les racines s'allongent. Il s'agit d'une photomorphose spéciale, provoquée ici par la plus ou moins grande intensité de la lumière latérale. Mais les mêmes effets sont observés dans le milieu forestier, ou bien en chambre de climatisation, sur des sujets germant dans une ombre générale plus ou moins dense, ou au contraire, en pleine lumière. Les conséquences culturales sont importantes car, de la forme des plantules, réglée ici par la seule lumière, dépendent leurs exigences en eau et en matières minérales, leur sensibilité éventuelle à certaines substances phytotoxiques, etc.... dont la distribution n'est pas homogène dans les couches superficielles du sol. Et ceci constitue, sans doute, une justification " a posteriori " de la vieille pratique culturale des coupes d'ensemencement.

Chênes pédonculés âgés de 3
ans. À gauche, sujet dont la tige a été ombragée
par un, puis par deux manchons de poterie, atteignant 0,90 m de longueur totale.
À droite, sujet à tige non ombragée, atteignant à
peine 0,25 m de hauteur totale.
On réalise assez bien ce qui peut se passer : la lumière,
pénétrant à l'intérieur des tissus, vient, d'une
façon générale, inactiver partiellement l'AIA, soit au
moment de son transfert, soit à l'endroit où il agit. Si, par
suite de l'ombragement latéral de la tige, la lumière interne
diminue, la concentration des tissus en AIA augmente. On peut ainsi passer,
en théorie, sur la figure 1, d'une concentration de 10-8 (sujets
à tige bien éclairée, s'allongeant modérément,
et à rameaux latéraux assez développés), à
une concentration de 10-6 à 10-5, par exemple (sujets
à tige ombragée, nettement plus longue, alors que les rameaux
latéraux sont réduits). Le schéma de THIMANN est donc encore
valable ici. De toutes façons, cet " effet manchon " doit
être rencontré fréquemment dans le milieu forestier, son
étude n'est qu'à peine commencée, mais ses incidences culturales
sont certainement nombreuses.
Cas des arbres développés en peuplements denses, et dont les troncs sont brusquement mis en lumière.
Il est d'observation courante, dans le Nord-Est de la France, que les arbres
de certaines espèces (chênes et charmes notamment), développés
en peuplements denses, ont des troncs nets et à peu près dépourvus
de branches latérales ; ceci, sur une bonne partie de la hauteur de leurs
fûts. Si, à la suite d'une opération culturale d'intensité
un peu marquée : coupe des taillis dans un taillis sous futaie à
réserves peu nombreuses, éclaircie très forte dans une
jeune futaie, mise en lumière de plantations résineuses installées
dans des peuplements feuillus, etc... l'intensité de l'éclairement
des troncs vient à être sensiblement majorée, on ne constate
pas d'effets bien marqués dans l'été qui suit l'opération.
Mais, dès la seconde année, et de plus en plus au fur et à
mesure que les années s'écoulent, on observe que les troncs des
arbres ainsi isolés se couvrent de petits rameaux latéraux, qui
deviennent ensuite de petites branches (gourmands) et dont la présence
peut déprécier la qualité du bois formé ultérieurement.
CATINOT (9) fait la même remarque dans les massifs équatoriaux.
On peut penser à un réchauffement local des écorces, par
les radiations solaires qui favoriseraient ainsi la production des gourmands.
Mais on n'a pas de données précises, d'expériences concluantes,
pour étayer cette hypothèse. Par contre, on peut être assuré
que, par suite de cette opération culturale, la quantité de lumière
qui atteint la surface extérieure des écorces, est fortement majorée.
Des mesures instantanées faites au luxmètre, dans divers peuplements,
et par des types de temps variés, permettent de fixer cette majoration
de l'éclairement : en moyenne de 25 à 50 lux, dans un peuplement
très dense, il peut passer à 1.000 ou 2.000 lux, dans un peuplement
moyennement ouvert, et à 4.000 ou 5.000 lux, dans un peuplement très
clair. Il s'agit, bien entendu, de la valeur de l'éclairement à
direction horizontale.
Comme la densité optique des écorces, bien que très élevée,
n'est nullement infinie (on peut mettre en évidence des facteurs de transmission
optique de l'ordre de 1/1.000.000), on doit admettre que l'assise cambiale reçoit,
chaque seconde, un flux de 109 photons de lumière visible,
par centimètre carré. On constate donc un processus inverse de
celui décrit au paragraphe précédent sur " l'effet
manchon ". À la suite du brusque éclairement des troncs,
la teneur en AIA des tissus voisins du cambium diminue, et l'on repasse de la
concentration favorisant la croissance de la tige à celle stimulant la
croissance des bourgeons, lesquels se développent en gourmands. La plupart
des auteurs admettent que ceux-ci ont une origine proventive (GUINIER, PERRIN,
VENET). Par ailleurs, les gourmands donnent naissance à de petites branches
qui détournent à leur profit une partie de l'eau et des substances
minérales absorbées par les racines ; elles peuvent alors se développer
considérablement et provoquer l'arrêt de la croissance des parties
supérieures (descentes de cimes).
Ainsi donc, le schéma de THIMANN est encore valable, et il permet, sinon
de comprendre parfaitement, tout au moins d'englober dans un système
de représentation cohérent, un certain nombre de morphoses dues
probablement à l'intervention simultanée de l'AIA et de la lumière.
CONCLUSIONS
On peut tirer des quelques considérations qui précèdent
de nombreux commentaires, dont certains sont déjà venus, probablement,
à l'esprit du lecteur ; un point important est sans doute le suivant
:
En photologie forestière, on mesure, en général, la composante
verticale de l'ensemble du rayonnement naturel (ou des seules radiations visibles).
Ceci, pour travailler en parfait accord avec les météorologistes,
lesquels considèrent surtout l'éclairement énergétique
- (ou lumineux) - reçu sur une surface horizontale. Il faut reconnaître
le bien-fondé de cette position, en remarquant, en outre, que les feuilles
étant assez souvent dans une position pas très éloignée
de l'horizontale, l'intensité de la lumière verticale conditionne
la marche de la photosynthèse. Mais, ce faisant, on néglige une
partie importante du rayonnement circumglobal : celui dont la composante horizontale
semble, au niveau des tiges, ou au travers des écorces, agir d'une façon
parfois très marquée sur la morphogenèse (phototropisme,
allongement des tiges, développement des rameaux latéraux). Il
sera donc utile, dans le milieu forestier, de procéder à des mesures
de l'éclairement à direction horizontale, tout aussi bien que
de celui à direction verticale.
Passant de la méthodologie à la physiologie, on ne peut, avec
KOPFER (5), que s'étonner " qu'une molécule aussi simple
que celle de l'AIA ait, dans la cellule, un sort aussi mal connu ". Ceci
tient, selon cet auteur, " aux nombreux systèmes régulateurs
qui jouent autour de l'AIA, et contribuent à l'équilibre naturel
de la croissance ; ils empêchent d'isoler son action, et augmentent la
difficulté des recherches ". On doit évidemment souhaiter
que ces travaux, d'un intérêt fondamental, soient poursuivis dans
de nombreux laboratoires, et, de préférence, sur des espèces
forestières économiquement intéressantes.
Enfin, d'un point de vue sylvicole plus pratique, on doit souligner l'intérêt qu'il y aurait à procéder à des recherches photologiques dans le sens indiqué par CATINOT (9) : quelles espèces tropicales sont sensibles au phototropisme ? à l'éclairement circumlatéral ? quelles sont les variations des caractères morphologiques, anatomiques et technologiques des sujets selon les traitements appliqués ? Travail de longue haleine, certainement difficile, mais exaltant et probablement fructueux...
BIBLIOGRAPHIE SUCCINCTE
1. ROUSSEL (L.) 1973. -Un nouveau venu en physiologie végétale : le phytochrome, Bois et Forêts des Tropiques No 149, p. 53-57.
2. RICARD (J.) 1960. - La croissance des végétaux, Presses Universitaires de France, 128 p.
3. JACQUIOT (C.) 1970. - La Forêt, Masson, 170 p.
4. THIMANN (K. V.) et CHAMPAGNAT (P.) 1955. - Colloque consacré à la croissance des végétaux et aux corrélations de croissance, Bulletin de la Société Française de Physiologie végétale No 1, p. 7-23.
5. KOPFER (L.) 1969. - Croissance et développement des plantes, Gauthier Villars, 234 p.
6. ENGLER (A.) 1924. - Heliotropismus und Geotropismus der Baüme und deren waldbaüliche Bedeutung. Mitteilungen der schweizerischen Centralanstalt für das forstliche Versuchswesen, Zurich.
7. TRONCHET (A.) 1972. - Les dispositifs sensoriels des végétaux supérieurs, Institut Botanique de Besançon, 97 p.
8. BRIGGS (W. B.) 1964. - Phototropism in higher plants (In Photophysiology, Vol. 1), Academic Press, p. 223-271.
9. CATINOT (B.) 1969. - Les éclaircies dans les peuplements artificiels de la forêt dense africaine, Bois et Forêts des Tropiques No 126, p. 15-38.
10. ROUSSEL (L.) 1972. - Contribution à l'étude de l'effet manchon. Annales scientifiques de l'Institut Botanique de Besançon, 3e série Botanique, Fasc. 12, p. 15-22.
N. B. On trouvera, en outre, de nombreux développements sur le sujet traité, dans un important article de CHARTIER P., BECKER M., BONHOMME B. et BONY J. P. : " Effets physiologiques et caractérisation du rayonnement solaire dans le cadre d'une méthode d'aménagement sylvicole en forêt dense africaine." Bois et Forêts des Tropiques (1973), No 152, pp. 19-35.