Contribution à l'étude de l '"effet manchon"
par Louis Roussel
Article publié en 1972 dans les Annales Scientifiques de l'Université de Besançon, 3° série - Botanique, fasc. 12 - (version .pdf - 422 k)
|
Contribution à l'étude de l '"effet manchon" par Louis Roussel |
|
| Chênes pédonculés âgés de 3 ans. À gauche, sujet dont la tige a été ombragée par un, puis par deux manchons de poterie, atteignant 0,90 m de longueur totale. À droite, sujet à tige non ombragée, atteignant à peine 0,25 m de hauteur totale. |
Résumé. - Il résulte des mesures effectuées sur Quercus pedunculata et Crataegus monogyna que la protection latérale des tiges semble favoriser la multiplication des cellules et non leur élongation. L'auteur propose différentes hypothèses pour expliquer ce phénomène qu'il appelle " effet manchon ".
INTRODUCTION
L'étude systématique de l'influence globale du rayonnement naturel
sur le développement des espèces ligneuses installées sous
des couverts divers, dans le milieu forestier, fait apparaître très
rapidement un certain nombre d'anomalies.
Certes, on peut admettre à priori, que la croissance des jeunes arbres
est déterminée, principalement, par l'activité photosynthétique
de leurs appareils foliacés (aiguilles ou feuilles). En effet, des mesures
précises effectuées grâce aux instruments modernes à
absorption de rayonnement infrarouge montrent que, dans des conditions normales
d'alimentation en eau du sol, et avec une atmosphère contenant une proportion
de gaz carbonique voisine de 0,03 %, la quantité de ce gaz absorbé
croît avec l'intensité de la lumière incidente : d'abord
très vite, dès que l'on a dépassé le point de compensation,
puis de plus en plus lentement, pour aboutir à une allure asymptotique
finale, à un palier de saturation lumineuse, variant de 20.000 à
30.000 ou 50.000 lux, en général. Si l'on considère qu'en
plein découvert, dans le nord-est de la France, l'éclairement
moyen annuel peut être évalué à 27.000 lux environ,
on peut en déduire que si l'eau du sol ne fait pas défaut, le
développement des jeunes végétaux ligneux est directement
conditionné par l'importance du couvert dominant les diverses stations
forestières. L'optimum est atteint quand ce couvert a été
considérablement réduit, ou même a complètement disparu.
C'est ainsi que, dès l'année 1952, l'auteur, à l'occasion
de la thèse de Docteur Ingénieur qu'il avait soutenue devant la
Faculté des Sciences de Besançon, avait signalé l'existence
d'une corrélation très nette, dans le Haut-Doubs, chez Picea
excelsa, entre la valeur du rayonnement relatif (Rr), ou de l'éclairement
relatif (Er) reçu dans les diverses stations, et le développement
général des sujets, la longueur et le diamètre de leur
pousse annuelle, l'importance de leur appareil radiculaire, etc... Ceci, jusqu'à
un Rr d'environ 50 %, et pour des sujets âgés en général
de 1 à 15 ou 20 ans (ROUSSEL, 1953).
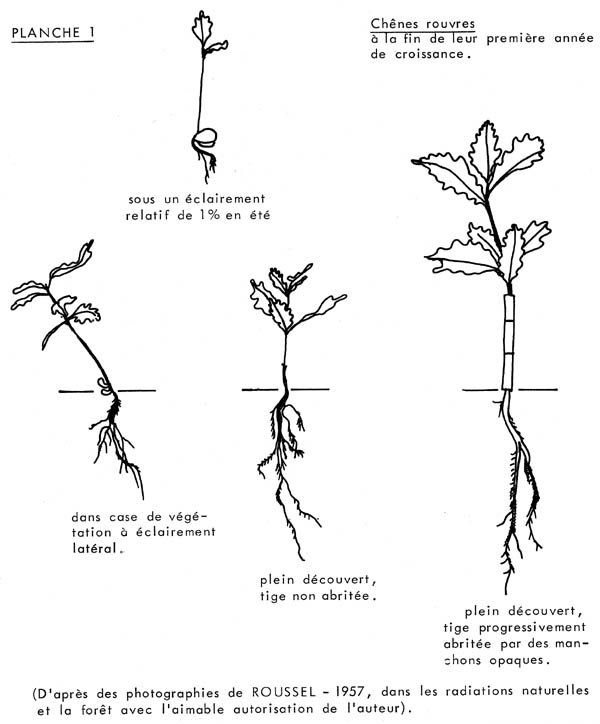
En recommençant ces expériences en cases de végétation à sol identique, dans la même région, l'auteur avait relevé les mêmes différences de comportement entre des sujets appartenant à 6 espèces résineuses (Coniférales), développés sous des Rr de 6 %, 12 %, 36 % et 100 %, et ce, dès leurs premières années de croissance. En particulier, pour Abies pectinata, considéré généralement comme une " essence d'ombre " typique, on ne constatait pas que les sujets ayant crû sous 6 % et 12 % de Rr étaient plus développés, à l'âge de 2 ans, que ceux qui avaient poussé sous 36 % ou sous 100 % de Rr. Au contraire, aux faibles intensités de rayonnement relatif, les sujets d'une longueur hors du sol très voisine étaient plus grêles, leurs appareils radiculaires moins fournis, en somme dans un état moins satisfaisant que ceux développés sous un Rr modéré, ou élevé (ROUSSEL, 1955).
1. COMPORTEMENT DU QUERCUS PEDUNCULATA ET DU QUERCUS SESSILIFLORA
Quelques années après les premières publications de l'auteur,
A. TRONCHET et A. GRANDGIRARD (1956), analysant de jeunes sujets de Quercus
pedunculata, âgés d'une année, et développés
sous des Er de 2 %, 15 %, 50 % et 100 0/, dans des expériences
organisées par G. PLAISANCE en forêt de Chaux, trouvaient que,
si le poids moyen, le diamètre de la tige et celui de la racine étaient
plus importants pour les sujets bien éclairés que pour ceux qui
avaient crû à l'ombre, la longueur de la tige par contre était
plus élevée dans les stations ombragées. G. PLAISANCE ne
constatait pas du reste que sous un Er de 2 %, considéré
comme très bas dans le milieu naturel, les jeunes chênes, à
l'âge d'un an manifestaient une nette tendance au dépérissement
(GRANDGIRARD, 1954).
Cette observation ayant paru peu en accord avec l'opinion générale
des forestiers praticiens (qui considéraient Quercus pedunculata comme
une espèce réclamant très tôt la pleine lumière),
l'auteur a recommencé des expériences sur cette espèce
ligneuse (ainsi que sur Quercus sessiliflora), dans des cases de végétation
à sol identique, et sous 6 degrés de Rr : 3,5 % - 4,3 %
- 13,5 % - 57 % - 66 % et 100 % (en été), dans un taillis
sous futaie de la Haute-Saône.
Au bout d'un an, les conclusions étaient analogues à celles de
G. PLAISANCE : dans toutes les stations, même sous les intensités
lumineuses les plus basses, les sujets étaient en vie, bien verts et
d'apparence florissante. Il apparaissait cependant que l'importance des matières
de réserve du gland (d'un poids moyen voisin de 2,5 grammes) permettait
de comprendre cette large tolérance au couvert au cours de la première
année de croissance car, dans leur seconde année, les jeunes chênes
situés sous 3,5 % de Rr disparaissaient alors que sous
4,3 % de Rr ils se maintenaient en vie, anémiés du reste
et peu florissants, et qu'ils persistaient dans cet état pendant quelques
années. Le point de compensation pouvait donc être fixé
dans ce cas à 4 % de Rr, seuil minimal de survie pour une glandée
normale (ROUSSEL, 1958). Cette conclusion a été vérifiée
par la suite, d'une façon très satisfaisante, à l'occasion
de deux glandées abondantes dans des taillis sous futaie de la Champagne
humide.
Les chênes de 2 ans, provenant de l'expérience décrite
ci-dessus, ont été analysés avec beaucoup de soin et de
manière approfondie à l'Institut Botanique de Besançon
(GIBOUDEAU, 1958; COGUELY, 1958, A. TRONCHET, J. TRONCHET, A.-M. GIBOUDEAU et
J. GOGUELY, 1959) et il est apparu alors un fait assez surprenant : un
certain nombre de caractères morphologiques ou histologiques des sujets
croissant sous des Rr relativement bas étaient différents
de ceux croissant en plein découvert. Par exemple, chez Quercus sessiliflora,
la longueur de la tige hors du sol était de 295 mm sous 13,5 %
de Rr et de 175 mm sous 100 % de Rr; la même
observation était faite pour la longueur de la racine 236 mm contre 212
mm, pour le volume de la tige 640 mm3 contre 430 mm3, etc...
Chez Quercus pedunculata, la longueur de la tige était de 284 mm sous 13,5 % de Rr et de 239 mm sous 100 % de Rr, la longueur de la racine de 240 mm sous 57 % de Rr contre 239 mm sous 100 %o de Rr, le volume de la tige de 1.000 mm3 sous 13,5 % de Rr, contre 730 mm3 sous 100 % de Rr. Que devenaient alors les vieilles notions forestières concernant les chênes, essences de " pleine lumière " ? Le plein découvert leur était moins favorable qu'aux jeunes sapins pectinés, pourtant réputés être des " essences d'ombre " bien caractérisées.
Hypothèse explicative
Devant ce comportement imprévu, l'auteur utilisant d'abord les données
histométriques établies dans les travaux précédemment
cités a proposé d'admettre que si la lumière à direction
verticale était incontestablement favorable à la nutrition carbonée
des sujets en expérience, il était possible que, chez les chênes,
la lumière horizontale venant frapper de tous côtés les
tiges en voie d'élongation et pénétrant au travers des
tissus de protection (épiderme avec sa cuticule, suber, fibres péricycliques,
etc...) vienne freiner la multiplication et l'élongation cellulaires
- un peu comme une lumière unilatérale, en cases de végétation
ouvertes d'un seul côté, provoque la classique courbure phototropique
de nombreux végétaux. Des essais de détermination de la
densité optique de ces formations de protection (grâce au microdensitomètre
très précis du CNRF de Nancy) montraient du reste l'exactitude
des résultats obtenus quelque temps auparavant par la technique plus
rudimentaire des photogrammes (fragments d'écorces appliqués directement
sur un papier photographique lent) : les formations de protection ne sont pas
absolument opaques et, dans le cas des jeunes chênes développés
en pleine lumière, leur densité optique oscille entre 3 et 4 (ce
qui correspond à un facteur de transmission optique de 1/1.000 à
1/10.000). Ayant établi par des mesures répétées
au luxmètre que la composante horizontale de la lumière, à
plein découvert, représentait de 4 à 5.000 lux, en moyenne,
l'auteur arrivait ainsi à déterminer un éclairement interne,
au niveau du cambium, de l'ordre de 0,5 à 5 lux. C'est cette faible lumière
qui pouvait donc intervenir pour ralentir l'élongation.
L'hypothèse était assez facile à vérifier : chez
des chênes dès leur germination ou au moment de leur croissance
annuelle il fallait, tout en laissant le méristème apical puis
les feuilles sous une lumière à composante verticale aussi élevée
que possible, protéger (par des manchons plus ou moins étroits),
la tige seule de la composante horizontale de cette même lumière.
Ce faisant, du reste, on se rapprochait des conditions de développement
des chênes dans le, milieu forestier, souvent affecté de trouées
de formes diverses. La lumière y est alors dispensée surtout verticalement,
alors que la lumière à direction horizontale est considérablement
réduite.
Ce sont ces expériences qui ont été relatées par l'auteur dans deux cour tes publications parues dans les Annales Scientifiques de l'Université de Besançon (ROUSSEL, 1962 et 1963). Pour fixer les idées, voici quelques-uns des résultats obtenus; les différences observées sont statistiquement significatives.
A) PROTECTION PAR DE PETITS MANCHONS OPAQUES APPLIQUÉS SUCCESSIVEMENT
AUTOUR DES TIGES, DÈS LA GERMINATION, ET LAISSANT TOUJOURS LES MÉRISTÈMES
APICAUX, PUIS LES FEUILLAGES, EN PLEINE LUMIÈRE :
|
Espèces
|
Quercus sessiliflora
|
Quercus Pedunculata
|
||
Conditions expérimentales
|
Sujets non protégés
latéralement
|
Sujets protégés latéralement
|
Sujets non protégés
latéralement
|
Sujets protégés latéralement
|
|
Longueur moyenne de la tige à
1 an....
|
10,1 cm
|
16,6 cm
|
13,4 cm
|
22,7 cm
|
|
Longueur moyenne de la tige à
2 ans...
|
13,4 cm
|
28,5 cm
|
20,3 cm
|
39 cm
|
B) PROTECTION PAR UN MANCHON LARGE DE POTERIE (HAUTEUR DE 37 CM ET LARGEUR DE
15 CM) DE SUJETS DE Quercus pedunculata, DÉJÀ AGÉS
D'UN AN, DÉVELOPPÉS JUSQU'ALORS DANS UNE TRÈS LARGE CLAIRIÈRE
:
|
Conditions expérimentales
|
Sujets non protégés
latéralement
|
Sujets protégés latéralement
|
|
Longueur moyenne du segment de tige
formé l'année de l'expérience...
|
9,3 cm
|
19,9 cm
|
|
Longueur moyenne totale de la tige
au bout d'un an d'expérience...
|
20,8 cm
|
32,1 cm
|
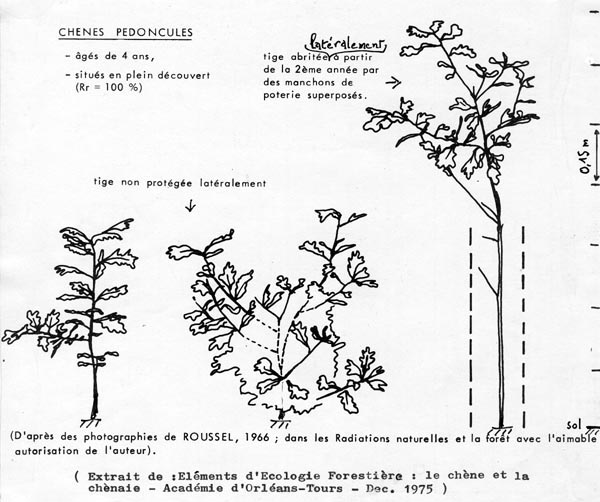
Cette dernière expérience, effectuée sur Quercus pedunculata, a été prolongée par adjonction d'un second manchon superposé au premier et de nouvelles séries de mesures ont été réalisées au bout de 3 années de protection (âge des sujets : 4 ans). Voici quelques-uns des chiffres relevés :
|
Conditions expérimentales
|
Sujets non protégés
latéralement
|
Sujets protégés latéralement
|
|
_
|
_
|
_
|
|
Hauteur totale au-dessus du sol
|
40 à 50 cm
|
90 à 110 cm
|
|
Développement des racines
|
important
|
assez réduit
|
|
Diamètre de la lige au collet
|
7,5 mm
|
6,5 mm
|
|
Volume du bois hors du sol
|
6 à 8 cm3
|
l0 à 12 cm3
|
|
Nombre de rayons ligneux au collet
:
|
||
|
petits
|
12-15
|
8-10
|
|
gros
|
6-8
|
très rares
|
|
Qualité du bois
|
plutôt dur et nerveux
|
plutôt «doux»
|
|
Longueur des cellules du parenchyme
ligneux |
40 a 60 µ
|
40 a 60 µ
|
|
Longueur des cellules du parenchyme
médullaire |
20 à 25 µ
|
20 à 25 µ
|
|
Hauteur des rayons ligneux
|
100 à 120 µ
|
100 à 110 µ
|
Il ressort en particulier de ces chiffres que l ' " effet manchon " a comme conséquences, outre l'allongement des tiges déjà signalé, la production d'un volume " tige " supérieur, d'une qualité technologique meilleure (prédominance du bois " doux "), mais constitué de cellules et de fibres d'une longueur analogue à celle des chênes témoins se développant a proximité immédiate. La protection latérale des tiges semble favoriser ici la multiplication du nombre des cellules, dans le sens de la longueur, et non leur élongation individuelle (ROUSSEL, 1967).
Il est difficile pour des raisons pratiques de suivre pendant un grand nombre d'années le développement de chênes protégés artificiellement ou non protégés. Mais il est possible d'observer parfois dans le milieu forestier des photomorphoses d'un genre analogue qui, bien que non réalisées à l'occasion d'expériences systématiques, peuvent cependant leur être assimilées quand on connaît exactement l'histoire des peuplements.
II. COMPORTEMENT DU CRATAEGUS MONOGYNA
En 1971, visitant dans le Haut-Doubs une plantation d'épicéas d'âge bien déterminé (20 ans) et d'une hauteur de 6 à 8 mètres, dense et bien venante, l'auteur a trouvé des sujets de Crataegus monogyna, installés naturellement peu après la mise en place des plants, dont certains se sont développés en dehors de cette formation, dans une haie naturelle de protection bien éclairée alors que d'autres, à l'intérieur de la plantation, croissaient et s'allongeaient en même temps que les résineux en maintenant leurs feuillages en pleine lumière. Dans la région inférieure de ce peuplement le Rr était, en 1971, de 0,5 %, donc très faible.
À l'âge maximum d'environ 20 ans, les sujets de Crataegus restés toujours bien éclairés sont courts (3 à 4 m de hauteur totale dont à peu près 1,50 m pour la tige principale), branchus, à cimes bien développées et à tiges épaisses, de 5 à 6 cm de diamètre à la base. Les feuilles sont assez petites et les épines abondantes.
Les seconds, d'un âge très voisin, abrités par l'ombrage des épicéas qui les enserraient, sont beaucoup plus longs (6 à 8 m de hauteur totale), ont des branches grêles, une cime plus réduite et rassemblée à la partie supérieure. Les tiges sont très nettement plus minces (3 à 4 cm de diamètre à la base) ; les feuilles sont plus larges et les épines peu abondantes. Par ailleurs, le bois produit dans ces deux sortes de sujets a été analysé avec beaucoup de soin à l'Institut du Bois (Laboratoire de C. JACQUIOT); voici les principaux résultats de ces mesures :
|
Conditions de croissance
|
Sujets non protégés
latéralement (courts)(d'environ 15 ans)
|
Sujets protégés latéralement
(longs)(d'environ 12 ans)
|
|
_
|
_
|
_
|
|
longueur moyenne des fibres
|
749 à 908 µ
|
827 à 869 µ
|
|
Diamètre moyen des fibres
|
16,6 à 17 µ
|
16,4 à 16,6 µ
|
|
Longueur moyenne des éléments
de vaisseaux
|
260 à 307 µ
|
301 à 322 µ
|
|
Nombre moyen de vaisseaux par mm2
|
180 à 235
|
179 à 239
|
|
Hauteur moyenne des rayons ligneux
|
231 à 233 µ
|
198 à 262 µ
|
|
Nombre moyen des cellules, en hauteur,
dans les rayons ligneux
|
10 à 12
|
12 à 13
|
N.B. - Ces nombres représentent les moyennes de nombreuses mesures ; certaines portent sur 50 à 100 éléments et sont effectuées dans chaque cas sur des segments homologues (base et région moyenne) des tiges des sujets " courts " et des sujets " longs ". Les moyennes Indiquées, soit 749 à 908 µ, par exemple, indiquent que ces moyennes varient, selon que l'on a opéré sur des segments prélevés à la base, ou au milieu de la tige. Mais ces chiffres présentés du plus faible au plus élevé ne signifient pas que les plus réduits correspondent à la région basse, et les plus élevés à la région moyenne.
En somme, et c'est là un résultat très intéressant, on retrouve chez Crataegus monogyna le même type de phénomène que celui observé chez Quercus pedunculata : l'ombragement latéral des tiges a favorisé très nettement la multiplication du nombre des cellules (mérèse) dans le sens de la longueur et pas du tout l'augmentation de leurs dimensions individuelles (auxèse) (ROUSSEL, 1972).
III. ESSAI D'INTERPRÉTATION PHYSIOLOGIQUE DE L' " EFFET MANCHON
"
Les résultats qui viennent d'être exposés ne surprendront
pas les botanistes qui d'une façon générale admettent que
" la lumière ralentit la croissance " et que, a contrario,
l'ombrage la favorise. Cependant, afin de tenter une analyse plus complète
de l' " effet manchon ", les expériences complémentaires
suivantes ont été effectuées sur Quercus pedunculata,
Quercus sessiliflora et Quercus rubra.
A) CROISSANCE DE SUJETS COMMENÇANT À GERMER, PLACÉS ENTIÈREMENT
EN OBSCURITÉ COMPLÈTE ET CONTINUE PENDANT 3 MOIS :
Dans ce cas on obtient des sujets à tige grêle, très longue
(de 30 à 40 cm), décolorée, et à feuilles réduites
à l'état de petites écailles, en somme typiquement "
étiolés ". La mort survient quand les réserves du
gland sont entièrement épuisées.
B) CROISSANCE DE SUJETS, COMMENÇANT À GERMER, EN PLEIN DÉCOUVERT,
DONT LES MÉRISTÈMES APICAUX SONT RECOUVERTS D'UN PETIT CAPUCHON
NOIR OPAQUE (D'UNE HAUTEUR D'ENVIRON 3 A 4 CM) :
Dans 80 % des cas, les chênes munis d'un capuchon apical arrêtent
la croissance en longueur de leur tige à un niveau très voisin
de celui des chênes témoins. Une ou deux feuilles se développent
alors latéralement, leurs pétioles étant recourbés
sous les capuchons, et prennent des dimensions et un aspect normal. Cependant,
dans 20 % des cas l'élongation se poursuit jusqu'à une
hauteur double de celle des témoins.
C) CROISSANCE DE SUJETS ABRITÉS LATÉRALEMENT SUR UNE HAUTEUR
ÉGALE À CELLE DES TÉMOINS, LES MÉRISTÈMES
APICAUX RESTANT BIEN ÉCLAIRÉS :
On obtient ainsi des sujets qui atteignent, au total, une hauteur double de
celle des témoins. Le déclenchement des processus qui règlent
la croissance en longueur de la tige, puis le développement des premières
feuilles, paraît donc se produire non à partir du niveau du gland
mais à partir de l'endroit où les méristèmes terminaux
apparaissent en pleine lumière.
Ce type d'expérience fait penser à une lente destruction, par
la lumière, d'une substance de croissance qui monte du gland ou des tissus
formés l'année précédente avec les éléments
plastiques indispensables à l'élaboration de la tige et des premières
feuilles.
D) MODIFICATION DU POUVOIR DE RÉFLEXION SPECTRAL DES ÉCORCES :
L'état lumineux qui règne à l'intérieur des tissus
des jeunes chênes dépend d'une part du pouvoir de réflexion
spectral des écorces, d'autre part du pouvoir d'absorption spectral des
formations de protection mentionnées plus haut. On a donc essayé
de modifier ce pouvoir de réflexion en appliquant des colorants non toxiques
sur les jeunes tiges de chênes au moment de leur croissance, afin de tenter
de déceler quelles radiations paraissent les plus efficaces pour ralentir
leur élongation. Parfois, les résultats sont très nets
et statistiquement significatifs : le noir, absorbant, toutes les radiations,
ralentit l'élongation ; le blanc et le bleu, qui ont un pouvoir
réflecteur marqué, favorisent au contraire l'allongement des tiges.
Mais dans d'autres cas, en revanche, on ne peut mettre en évidence aucun
effet.
Ces quatre séries d'expériences ont fait l'objet de communications
au Groupe d'étude de Physiologie de l'arbre (Président : C. JACQUIOT).
E) EXTENSION DE L'EFFET MANCHON AUX CONIFÉRALES :
Par ailleurs, on avait déjà recherché si l'allongement en abri circulaire latéral, observé chez les chênes, était également perceptible chez diverses espèces de Coniférales, indigènes ou exotiques, banales en France (une vingtaine au total). Pendant leurs deux premières années, toutes les espèces en expérience se sont révélées phototropiques ; elles s'allongent nettement en abri latéral. Mais dès leur troisième année en général, ces espèces perdent à la fois leur " faculté d'allongement " en abri latéral et leurs " propriétés phototropiques ". Ce genre d'expérience suggère - tout au moins en ce qui concerne les Coniférales - que les deux types de réaction ont la même origine physiologique (ROUSSEL, 1966).
Il est à remarquer à ce propos que J.F. NITSCH et L. SOMOGYI (1958), opérant sur de très nombreuses espèces ligneuses, et en particulier sur Picea excelsa au cours de sa troisième année de croissance et sur Pinus strobus au cours de ses cinquième et sixième années de croissance, ont montré que la modification expérimentale du régime photopériodique (faisant vraisemblablement intervenir, selon l'auteur, une interconversion des phytochromes) provoquait des changements très importants dans l'allure de l'élongation des sujets. Si les phytochromes intervenaient dans l' " effet manchon ", on comprend mal qu'ils cessent d'agir quand les jeunes Coniférales ont atteint l'âge de 2 ans, alors qu'ils continuent à se manifester dans les photomorphoses provoquées par la modification du régime photopériodique sur des sujets plus âgés.
IV. DISCUSSION
Quels types de processus, actuellement bien décrits, pourraient donc être invoqués pour expliquer cet " effet manchon " ?
PROCESSUS A
La lumière du jour, riche en rayons rouge clair, provoque chez des sujets de plein découvert une formation prédominante de phytochrome 730. Selon P. ROLLIN (1970), le rapport du phytochrome 730 au phytochrome total (P 730 + P 660) peut alors y atteindre 60 %. Ce phytochrome 730, sans doute en favorisant la photo-oxydation de l'acide ß indole-acétique, ralentit la croissance en longueur. Quand la tige est ombragée, le phytochrome 660 redevient prédominant ce qui permet l'allongement des tiges. Ceci, même en régime photopériodique constant.
Cependant, il faut admettre selon cette explication que la tige seule peut jouer le rôle de photorécepteur (alors que l'on pensait jusqu'ici que ce rôle était joué par les feuillages). Par ailleurs on comprend mal, on l'a dit, le comportement des Coniférales décrit au paragraphe E ci-dessus.
Mais l'allongement des tiges décrit dans l'expérience du paragraphe A (croissance des sujets en obscurité complète et continue) pourrait sans doute être rattaché à une intervention des phytochromes provoquant un étiolement typique.
PROCESSUS B
La lumière du jour déplace partiellement les substances de croissance de nature auxinique des parties éclairées vers les parties ombragées. C'est la première version de la théorie de CHOLODNY-WENT (in BRIGGS, 1964) qui explique très bien à elle seule l'effet phototropique. Mais comme selon cette théorie la quantité totale d'auxines reste la même (seule change la répartition dans la tige) on comprend mal l'élongation observée quand la tige seule des chênes est bien ombragée.
PROCESSUS C
C'est le processus décrit par C CHOLODNY-WENT, mais passablement modifiée par de nombreuses expériences ultérieures (celles de GALSTON et de BRIGGS notamment (in BRIGGS, 1964). Au déplacement d'une substance de croissance bien connue cette fois, l'acide ß indole-acétique, s'ajoute une photooxydation provoquée par la lumière bleue de cette même substance (la riboflavine jouant probablement le rôle de photosensibilisant).
Mais dans ce cas, il faut reconnaître que les détails de ce processus tels qu'ils ont été décrits chez le coléoptile d'Avoine (élaboration de l'acide ß indole-acétique à l'apex, transfert polarisé vers les régions inférieures, avec modification de sa concentration en cours de trajet sous l'effet de la lumière interne des tissus végétaux, et surtout allongement des cellules de la partie moyenne du coléoptile), ne sont pas exactement reproduits dans le cas des tiges de chêne; cela ressort des diverses expériences relatées plus haut.
Par ailleurs, on doit remarquer que des expériences du genre de celles
réalisées par MILLET (1971) mettent en évidence chez certains
végétaux, à côté de la croissance apicale,
une croissance intercalaire qui intervient d'une façon importante pour
déterminer l'élongation totale constatée.
PROCESSUS D
On doit faire entrer en ligne de compte les modifications des facultés
d'allongement et de multiplication des cellules elles-mêmes, suivant
qu'elles sont soumises à un éclairement interne plus ou moins
intense. Cet effet renforcerait celui décrit précédemment.
Il s'agit de la vieille théorie de BLAAUW, (in SAUBERER et HAERTEL,
1959), récemment mise à jour par GUTTENBERG, toujours du reste
sur le coléoptile d'Avoine.
CONCLUSION
La solution de facilité consisterait à souligner que l' "
effet manchon " ne semble explicable, de manière stricte, par aucun
des processus qui viennent d'être sommairement décrits et à
suggérer l'intervention d'un mode d'action nouveau, tout à fait
différent de ceux actuellement reconnus.
Cependant, il existe de nombreuses analogies entre cet effet et ce qui est
observé dans le cas du coléoptile d'Avoine (processus C et D tout
spécialement). Aussi l'auteur propose-t-il d'admettre, tout au moins
à titre provisoire, que l'on se trouve en présence d'un phénomène
d'un type assez classique dans lequel effectivement " la lumière
ralentit la croissance ".
Mais il faut reconnaître dans ce cas l'existence de certaines modalités
originales, notamment des suivantes :
- L'organe photorécepteur paraît être principalement la
tige elle-même (et non les feuilles, ou le méristème terminal).
- La photo-inhibition de la croissance semble s'opérer plus tôt,
soit lors d'un transfert ascendant de la substance oligodynamique concernée,
soit lors de la croissance intercalaire en longueur suivant la croissance apicale
(et non lors d'un transfert polarisé descendant de l'acide ß indole-acétique).
- L'ombragement latéral de la tige favorise surtout la mérèse (multiplication du nombre des cellules dans le sens de la longueur mais réduction de ce nombre dans le sens du diamètre) - et non l'auxèse (majoration des dimensions longitudinales individuelles de celles-ci).
BIBLIOGRAPHIE
BRIGS W.R., 1964. - Phototropism in higher plants. In Photophysiology,
vol. 1, Academic Press, 223-271.
GIBOUDEAU AM., 1958. - Comportement de plants de Quercus sessiliflora
L. (Chêne rouvre), âgés de deux ans en présence de
conditions différentes de radiation relative. Diplôme d'études
.Supérieures, 60 p.
GUOGUELY J., 1958. - Réponses de plants de deux ans de Quercus pedunculata
soumis à six éclairements relatifs différents. Diplôme
d'Etudes Supérieures, Besançon, 67 p.
GRANDGIRARD A., 1954. - Observations sur la structure anatomique de plantules
de Quercus pedunculata, cultivées sous des éclairements relatifs
différents. Diplôme d'Etudes Supérieures, Besançon,
64 p.
MILLET B., 1971. - Caractéristiques d'un rythme circadien de la croissance
chez la Fève. Bulletin du Groupe d'Etude des Rythmes Biologiques,
t. 3, n° 4, 79-83.
NITSCH J.P. et L. SOMOGYI, 1958. - Le photopériodisme des plantes
ligneuses. Ann. Soc. nation. Hort. Fr., 4° année, n°
16, 466-490.
ROUSSEL L., 1953. - Recherches théoriques et pratiques sur la répartition,
en quantité et en qualité de la lumière dans le milieu
forestier. Influence sur la végétation. Ann. Ec. nation. Eaux
Forêts, 13, fasc. 2, 295-400.
ROUSSEL L., 1955. - Essences d'ombre, essences de lumière. Bull.
Soc. forest. Fr. Comté, juin 1955, I-Il.
ROUSSEL L., 1958. - Exigences minimales en lumière des chênes
de la Haute-Saône. Bull. Soc. forest. Fr. Comté, mars 1958,
1-II.
ROUSSEL L., 1962. - Les chênes et leurs écorces.
Ann. scient. Univ. Besançon, 2° sér., Bot. 18, 141-148.
ROUSSEL L., 1963. - Effets obtenus en ombrageant
la tige du chêne pédonculé. Ann. scient. Univ. Besançon,
Bot. 19, 67-71.
ROUSSEL L., 1966. - Le phototropisme juvénile
des résineux. Rev. forest. fr., n° 11, 708-717.
ROUSSEL L., 1967. - De l'emploi des modèles
réduits en sylviculture. Rev. forest. fr., n° 12, 737-745.
ROUSSEL L., 1972. - Nouveaux exemples de photomorphoses en régime photopériodique
identique. Communication au Groupe d'Etude de Physiologie de l'Arbre, non publiée.
SAUBERER F. et 0. HAERTEL, 1959. - Pflanze und Strahlung. Geest &
Portig, 268 p.
TRONCHET A. et A. GRANDGIRARD, 1956. - L'analyse histométrique et son
application à l'Ecologie forestière. Ann. scient.. Univ. Besançon,
2° sér. Bol. 8, 3-30.
TRONCHET A., J. TRONCHET, A.M. GIBOUDEAU et J. GOGUELY, 1959. - Etude morphologique et histométrique de plants de Quercus sessiliflora et de Quercus pedunculata, âgés de deux ans et soumis à différents éclairements relatifs. 84° Congrès National des Sociétés Savantes, 425-431.
documents annexes :