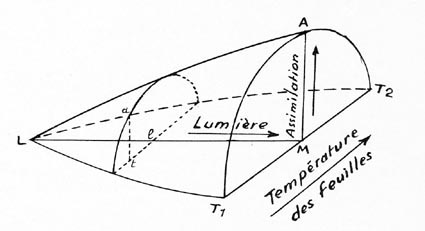
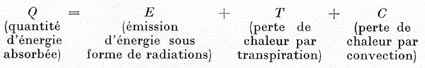Une situation microclimatique quelconque (lumière l, température
t de la figure 1) se traduit par une assimilation carbonée déterminée
: t. a. Quand il n'y a plus de lumière, en L, toute la photosynthèse
est arrêtée. Seule alors fonctionne la respiration (qui serait
représentée au-dessous du plan L-T1-T2 de la fig. 1).
La prise en considération de la température des feuillages est
donc, théoriquement, des plus importantes, aussi bien pour les sujets
situés en plein découvert que pour ceux qui se développent
dans les sous-bois.
On peut, très schématiquement, analyser de la façon suivante
la façon dont interviennent les multiples facteurs du milieu :
1) une feuille, détachée, placée sous un abri (du type
météorologique, par exemple) tend à se mettre à
une température égale à celle de l'air ambiant,
2) la même feuille, détachée, exposée aux rayonnements
solaire et céleste (dont elle repousse une partie grâce à
son albédo propre) tend à s'échauffer. Mais elle perd une
partie de cette chaleur par son rayonnement propre et par convection au contact
de l'air ambiant,
3) une feuille identique, mais reliée à un arbre vivant, outre
les processus décrits au paragraphe 2 ci-dessus, transpire, plus ou moins
activement, et tend maintenant à se refroidir, parfois nettement même
au-dessous de la température de l'air ambiant.
En outre, cette dernière feuille, fixe par la photosynthèse,
une proportion, généralement très faible du rayonnement
absorbé.
2. BILAN DES ÉCHANGES D'ÉNERGIE AU NIVEAU DE LA FEUILLE
L'équilibre entre l'absorption de l'énergie naturelle par la
feuille, et la déperdition de cette même énergie par cet
organe, peut être exprimé sous la forme simplifiée suivante
(en négligeant l'énergie fixée par la photosynthèse):
Voici quelques précisions sur ces divers éléments, tels
qu'ils résultent d'une importante étude de D. M. GATES et E. LAVERNE
PAPIAN (II) :
Q = Selon D. M. GATES, les feuillages absorbent environ 50 % de l'énergie
solaire directe et diffusée par le ciel, et environ 95 % de l'énergie
de grande longueur d'onde réémise par l'atmosphère
de la terre. Ceci donne, au total, les ordres de grandeur suivants, pour
une inclinaison des feuilles de 45 °C par rapport à ladirection
du soleil au milieu de la journée :
- Dans les déserts, en été, la quantité d'énergie
absorbéeest évaluée à 1,1-1,4 cal/cm2/mn.
- Dans les régions basses ou moyennes, de climat tempéré,
la même quantité d'énergie, en été, varie
de 0,95 à 1,1 cal/cm2/mn.
- Dans les régions alpines, en été, cette quantité
est de 0,8 à 0,95 cal/cm2/mn.
- En hiver, toutes ces valeurs diminuent (dans les régions alpines,
elles peuvent osciller entre 0,2 à 0,7 cal/cm2/mn.
E = Cette émission radiante, faible en général, est,
pour un feuillage déterminé, proportionnelle à la 4e
puissance de sa température absolue (loi de STEFAN-BOLTZMANN).
T = Cet élément est souvent le plus important, mais aussi
le plus complexe à évaluer, puisqu' interviennent, outre
naturellement la chaleur latente de vaporisation de l'eau transpirée,
la densité de vapeur saturante dans l'air et dans la feuille, l'humidité
relative de l'air, les dimensions de la feuille et la vitesse du vent, le
tout, en relation avec les températures respectives de l'air et de
la feuille.
C = La perte de chaleur par convection est conditionnée par la différence
de température entre l'air et la feuille, les dimensions de cette
feuille et la vitesse du vent.
Au total, D. M. GATES retient 9 variables :
La quantité d'énergie absorbée par la feuille, la vitesse
de l'air, la température de l'air, la température de la feuille,
la transpiration, l'humidité relative de l'air, la résistance
interne à la diffusion de l'eau, la dimension de la feuille dans le sens
du vent et la dimension de la feuille perpendiculairement au sens du vent.
Ces variables interviennent, dans une formule générale, avec
un certain nombre de constantes physiques classiques, ou résultant d'expériences
spéciales. Le calcul, dans chaque cas, est très compliqué.
Aussi, et c'est l'intérêt principal de l'ouvrage mentionné
(II), les principaux résultats sont présentés, pour des
valeurs d'encadrement caractéristiques, sous forme de tableaux (280)
et de graphiques (110) à 2 ou 4 variables. Il est ainsi facile de se
faire une idée globale de l'ensemble de ces phénomènes.
3. QUELQUES TYPES DE RÉSULTATS
Les régions désertiques tropicales.
Le climat désertique tropical, très chaud en été,
à faible pluviosité estivale, à air souvent sec, présente
des caractères très spéciaux, et, en général,
défavorables à la végétation. Toutefois, si les
végétaux peuvent trouver, parfois assez profondément dans
le sol, des réserves d'eau suffisantes, la transpiration peut aider efficacement
à refroidir les feuillages - donc à améliorer le rendement
de la photosynthèse, ainsi qu'on peut le voir ci-dessous :
Données générales :
Energie absorbée 1,433 cal/cm2/mn.
Résistance à la diffusion : 1 s/cm.
Température de l'air: + 40 °C.
Humidité relative de l'air: 0 % et 100 %
--- Vitesse du vent : 10 cm/s.
Température des feuilles de 1 cm x 1 cm = 35°,5 (HR 0 %) à
45°,4 (HR 100 %).
Température des feuilles de 20 cm x 10 cm = 44°,9 (HR 0 %) à
54 °C (HR 100 %).
--- Vitesse du vent : 100 cm/s.
Température des feuilles de 1 cm x 1 cm = 34°,8 (HR 0 %) à
44°,3 (HR 100 %).
Température des feuilles de 20 cm X 10 cm = 35°,9 (HR 0 %) à
47°,6 (HR 100 %).
NB. En conditions microclimatiques égales (sous réserve des variations
indiquées) et pour une surface foliaire totale égale : les petites
feuilles sont moins chaudes que les grandes feuilles la température des
feuilles est d'autant plus basse que l'humidité relative de l'air, même
chaud (40 °C) est plus faible, et que la vitesse de l'air est plus grande.
On note ici l'effet antithermique très net de la transpiration.
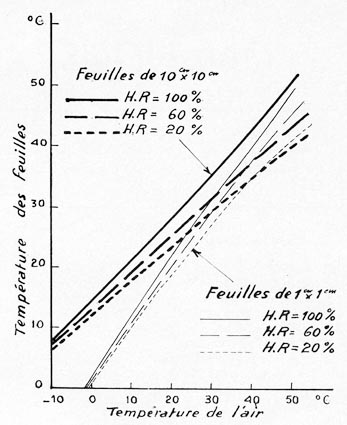
Radiation absorbée : 1,1 cal/cm2/sec
Résistance interne : 1 sec/cm
Vitesse du vent : 100 cm/sec
FiG. 2. - (Extraite des graphiques no 5 et 6 de D.
M. GATES et LAVERNE E. PAPIAN.)
Les régions basses tempérées en été.
En général, à une latitude voisine de 45°, le climat
estival, dans les régions basses, sous influence maritime, est caractérisé
par des journées à température assez élevée,
entrecoupées de jours plus frais, avec pluies orageuses, ou non, assez
abondantes. La végétation y trouve rarement des conditions climatiques
excessives aboutissant à des dommages marqués, ou à un
retard net de croissance. On retrouve, mais d'une façon atténuée,
les mêmes types de résultats que ci-dessus :
Données générales :
Énergie absorbée 1,15 cal/cm2/mn.
Résistance à la diffusion : 1 s/cm.
Température de l'air: + 30 °C.
Humidité relative de l'air : 0 % et 100 %
--- Vitesse du vent : 10 cm/s.
Température des feuilles de 1 cm x 1 cm = 27°,6 (HR 0 %) à
32°,8 (HR 100 %).
Température des feuilles de 20 cm x 10 cm = 29°,9 (HR 0 %) à
38°,6 (HR 100 %).
--- Vitesse du vent : 100 cm/S.
Température des feuilles de 1 cm x 1 cm = 28° (HR 0 %) à
32°,2 (HR 100 %).
Température des feuilles de 20 cm x 10 cm = 28°,3 (HR 0 %) à
37°,l (HR 100 %).
En réalité, les quelques valeurs reproduites ci-dessus ont été
choisies parmi les plus démonstratives et une analyse complète
des divers cas révèle bien plus de nuances et même fait
apparaître certaines inversions dans les conclusions générales
précédentes.
La figure 2 extraite de l'ouvrage analysé, montre comment se présentent
certains des 110 graphiques, plus généraux, mais moins précis,
que les 280 tables.
4. APPLICATIONS PRATIQUES
Il est légitime de se demander dans quelle mesure les résultats
ci-dessus, qui sont acquis, il faut bien le répéter, par une
méthode théorique (bien que basée sur des données
physiques bien classiques et, sur certains facteurs déterminés
expérimentalement), se raccordent avec des observations globales, faites
en laboratoire ou sur le terrain.
On peut citer quelques faits :
Les expériences de W. TRANQUILLINI (1964) relatées dans : Photologie
forestière, page 68
(I). Au phytotron du Patscherkofel, l'accélération de la transpiration
des aiguilles de jeunes mélèzes d'Europe peut abaisser leur température
de 10 °C environ, et, ainsi, majorer très sensiblement l'importance
de leur photosynthèse nette, en conditions microclimatiques par ailleurs
égales.
Les observations de H. TURNER (1971) dans les Haute Alpes de Suisse. Le développement
des mélèzes d'Europe et de l'épicéa commun, au voisinage
de 2.000 m d'altitude, est beaucoup plus important sur les versants ensoleillés
(lumière et chaleur plus fortes), que sur les versants ombragés.
Ceci par vent faible, inférieur ou voisin de 100 cm/s. Un vent plus fort
tend à égaliser la température du sol (et des aiguilles)
dans les diverses stations. Les différences de croissance s'atténuent
et disparaissent presque quand la vitesse du vent atteint 300 cm/s.
Le problème des sapinières de plaine peut, sans doute, être
compris en partie de cette façon. En altitude modérée (de
500 à 800 m par exemple) la température des aiguilles reste au
voisinage de celle qui correspond à l'assimilation carbonée optimale.
En plaine, si le sol est bien approvisionné en eau (cas de certaines
vallées par exemple), la transpiration contribue efficacement à
abaisser la température des aiguilles, dans une atmosphère généralement
plus chaude qu'en altitude, et il peut en résulter une croissance satisfaisante
de certains peuplements de sapins.
Dans les régions tropicales sèches, on rencontre fréquemment
des arbres et des arbustes à petites feuilles (du genre des mimosées,
par exemple). Cette morphologie fait, qu'en conditions microclimatiques identiques,
leurs cimes sont plus fraîches que celles des arbres à grandes
feuilles. Par ailleurs - mais il s'agit là d'un problème différent
- la constitution de plans successifs, en quinconce, pour les feuillages, permet
un rendement photosynthétique d'ensemble maximal, pour une surface foliaire
minimale.
Évidemment , il ne s'agit encore que d'observations isolées,
de concordances, apparentes peut-être, dues au hasard, ou bien à
l'intervention de processus encore ignorés des physiologistes. Il n'en
demeure pas moins que la question de l'équilibre thermique des feuillages
semble susceptible, pour les sylviculteurs, de développements intéressants.
BIBLIOGRAPHIE
- ROUSSEL, L. (1972). - Photologie
forestière. Masson.
- GATES, D. M. et LAVERNE PAPIAN, E. (1971). - Atlas of energy budgets of
plants leaves.Academic Press.