Article publié en 1978 dans la Revue Forestière Française, n° 3, p 186 - (version .pdf - 1,9 M)
Lumière,
gourmands et rejets de souche
| " Il est plus aisé de dire des
choses nouvelles, que de concilier celles qui ont été dites. " |
|
Vauvenargues.
|
Depuis très longtemps, les forestiers ont attaché une grande
importance au facteur " lumière ", d'influence très
complexe, certes, mais dans laquelle on a pu distinguer assez rapidement plusieurs
sortes d'effets, notamment :
- un effet stimulant sur la photosynthèse, donc sur la nutrition en
général (entre l'éclairement minimal de compensation et
l'éclairement maximal de saturation);
- un effet morphogénétique, ce terme comprenant des actions très
variées comme la courbure phototropique ou bien le modelage de l'architecture
même des arbres. L'auteur de cette analyse a évoqué certains
des aspects de ce vaste problème dans cette Revue même,
il y a quelques années déjà (1 - 2 - 3 - 4), (voir nota).
On a récemment avancé que la formation des gourmands, sur les
troncs des jeunes arbres mis en lumière à la suite d'une coupe
assez forte (sur de jeunes chênes, en particulier), pouvait être
provoquée par l'éclairement plus intense que reçoivent
alors, latéralement, ces troncs (5) ; de même certains auteurs
estiment que les rejets de souches, qui constituent la base même du traitement
en taillis, ont besoin pour apparaître et, bien entendu, pour se développer,
d'un apport de lumière, verticale cette fois (6 - 7). S'agit-il de processus
entièrement nouveaux, non encore étudiés par les physiologistes,
ou bien est-il possible d'interpréter ces faits grâce aux théories
modernes qui font appel à la physique, à la chimie, à la
physiologie végétale et, d'une façon plus précise,
à la photobiologie ? C'est ce qui va être examiné maintenant.
Nota : Les chiffres entre parenthèses renvoient à la bibliographie.
BREF RAPPEL DE QUELQUES PRINCIPES THÉORIQUES DE BASE
Le rayonnement solaire, direct et diffusé par le ciel, apporte la lumière
et la chaleur ; il est constitué de radiations dont la longueur
d'onde va, approximativement, de 300 nm (nm = nanomètre = 10-9 mètre),
limite de l'ultraviolet au sol, à 3000 nm environ, limite de l'infrarouge ;
cette plage est le domaine propre à la photobiologie qui s'intéresse
à ces radiations dites, on le verra plus loin, " non ionisantes
", Par contre, la radiobiologie s'attache à déterminer les
effets des radiations d'une longueur d'onde nettement inférieure à
300 nm : rayons X et y qui sont dits " ionisants " et qui peuvent
apporter des modifications profondes à la structure intime des être
vivants, souvent des destructions du reste ; au voisinage de 300 nm, on peut
observer les deux sortes d'effets qui se chevauchent partiellement.
Les radiations sont constituées de minuscules paquets d'énergie,
les photons, associés à des ondes électromagnétiques
; pour les radiations non ionisantes, l'énergie du photon varie approximativement
de 4 électrons-volts (*), à la limite de l'ultraviolet au sol,
jusqu'à 1,7 électron-volt, dans l'extrême rouge visible,
et à 0,4 électron-volt, pour les infrarouges les plus lointains.
(*) L'électron-volt (eV) est l'unité d'énergie à
l'échelle atomique et moléculaire ; c'est l'énergie cinétique
acquise par un électron soumis à une différence de potentiel
de un volt.
La matière est constituée, on le sait, de molécules formées d'atomes très stables, constitués chacun d'un noyau (proton[s] chargé[s] positivement, et neutron[s], électriquement neutre[s]) - et d'un ou de plusieurs cortèges d'électrons, chargés négativement, décrivant autour du noyau des orbites bien précises ; en fait, depuis de Broglie, on pense plutôt à des " orbitales ", zones de l'espace, sphériques, ellipsoïdales, etc. correspondant à des " probabilités de présence " desdits électrons. Tout en se déplaçant autour du noyau, les électrons tournent sur eux-mêmes et, comme ils sont chargés électriquement, il en résulte un moment magnétique qui varie avec leur sens de rotation : le " spin " est une grandeur liée à ce champ magnétique et cette notion joue un grand rôle dans tous les phénomènes photochimiques.
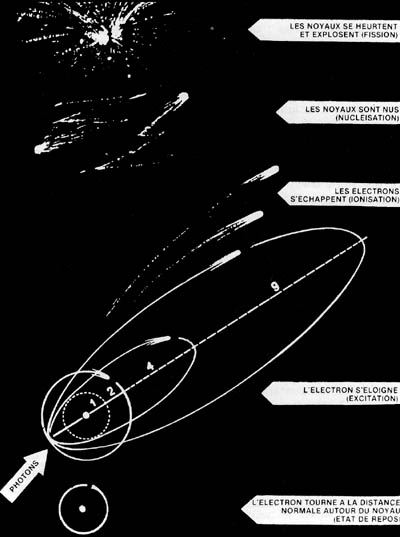
| Fig. 1 - Représentation schématique des effets des radiations sur la matière : sous l'action de photons peu énergiques (1 eV, vers le milieu de l'infrarouge) les orbitales des électrons ne sont pas modifiées (cercle 1, en pointillés). L'atome reste, dans l'exemple envisagé ici, à l'état de repos, ou " fondamental ". Quand l'énergie des photons atteint 2 eV (sensation visuelle de couleur vert-jaune), un électron passe sur une autre orbitale sphérique (cercle 2, en traits pleins) et l'atome prend un état dit " excité ". Le retour de l'électron sur son orbitale primitive est à l'origine de l'effet photobiologique. Avec des photons d'énergie croissante (de 4 à 9 eV) les orbitales sont de plus en plus allongées, et, à partir de 10 eV (limite de l'ultraviolet naturel en dehors de l'atmosphère terrestre), un électron est éjecté, et l'atome devient " ionisé ". Cette barrière d'énergie, variant avec la nature des substances, sépare, en théorie, la photobiologie de la radiobiologie. | |
Lorsque plusieurs atomes se réunissent pour former une molécule,
leurs noyaux restent séparés, mais les orbitales des électrons
se fondent partiellement en orbitales moléculaires, classées en
"orbitales sigma" relativement stables et assurant souvent la cohésion
des molécules et en " orbitales ì ", en général
plus fragiles. Pour arracher un électron à une orbitale (la molécule
devient alors " ionisée "), il faut que les photons possèdent
une énergie variant, en général, de 3 à 20 électrons-volts
; la lumière visible ne peut donc, la plupart du temps, produire cet
effet important. Mais, d'autres actions sont possibles : augmentation de l'agitation
thermique de la molécule (donc de sa température), déplacement
d'un électron de son orbitale habituelle (état fondamental) sur
une orbitale plus éloignée (état dit " excité
"). Cet effet étant obtenu avec des photons dont l'énergie
est " de l'ordre " de l'électron-volt, la lumière visible
peut donc fréquemment intervenir ; outre le déplacement de l'électron,
il peut se faire que son " spin " soit modifié, donc l'état
de l'ensemble de la molécule. Quand l'électron déplacé
reprend sa place primitive, il libère tout ou partie de l'énergie
qu'il avait reçue et il en résulte des effets variés :
émission longue ou brève de rayons lumineux, fragmentation de
la molécule, transfert d'énergie à d'autres molécules,
etc. Tout ceci est le domaine de la photochimie (concernant les substances minérales,
comme les substances organiques).
Un aspect spécialement intéressant de ce genre d'études
est l'intervention des substances photosensibilisantes : un corps insensible
à toute modification quand il est soumis seul aux rayons lumineux peut
voir ses propriétés modifiées très profondément
quand il se trouve à proximité d'une substance photosensibilisante
(presque toujours colorée*) qui capte l'énergie de certains photons
et la transfère sur lui. L'un des premiers exemples de ce mode d'action
a été réalisé par Raab, à Heidelberg, en
1898, quand il montra que des colonies de protozoaires, insensibles à
la lumière du jour, étaient détruites quand on ajoutait
à leur milieu un colorant, l'acridine, par ailleurs sans action sur eux
à l'obscurité.
(*) D'une façon générale, pour qu'une substance soit sensible à l'action de la lumière, il faut qu'elle absorbe certains groupes de radiations bien déterminés ; c'est l'une des lois de la photochimie " le spectre d'absorption coïncide avec le spectre d'action ". Il faut cependant prendre garde au fait que l'œil humain est insensible à certaines radiations, émises de façon permanente par le soleil et par le ciel.
Tout ce qui vient d'être dit sur les rapports de la lumière et de la matière revêt une importance considérable : la photosynthèse, base de toute vie sur notre globe, la vision également, ainsi qu'une foule d'autres effets auxquels on prête parfois peu d'attention, peuvent être interprétés grâce à ce corps de théories qui offre un très bon exemple d'accord entre le modèle et la réalité. Des ouvrages récents comportent du reste d'excellentes mises au point sur ce sujet (8-9-10).
**
Pour en revenir à l'intervention des photosensibilisateurs (effet dit " photodynamique ") et au sujet examiné ici, on doit apporter une attention spéciale aux photo-oxydations : en présence de substances intermédiaires très actives comme les acridines, azines, porphyrines, et surtout la riboflavine, existant chez de nombreux végétaux, et grâce à l'intervention de l'oxygène, diverses molécules organiques sont détruites par la lumière. On peut citer dans cet ordre d'idées la photo-oxydation du tryptophane, acide aminé très voisin de l'auxine naturelle (acide ß indole acétique ou par abréviation AIA) considéré généralement comme son précurseur, qui est transformé en N-formylkynurénine, et la photo-oxydation de l'auxine naturelle elle-même qui donne une aldéhyde, du gaz carbonique et de l'eau. Or, l'auxine naturelle ou AIA est l'une des hormones végétales la plus anciennement connue ; découverte par Went, vers 1928, elle fut isolée par Kôgl en 1934. Depuis cette époque, on a montré qu'elle intervenait très activement dans le développement des végétaux : allongement (auxèse) et multiplication (mérèse) des cellules, phototropisme et géotropisme, organisation des tissus, chute des feuilles et des fruits, etc. ; produite en permanence par les plantes, agissant à dose infinitésimale (les concentrations actives vont, suivant la nature des tissus, de 10-3 à 10-12), cette hormone devrait s'accumuler chez les végétaux, s'il n'existait des mécanismes de régulation, comme la synthèse de l'auxine-oxydase, enzyme qui réduit en permanence son taux de concentration. Mais l'auxine-oxydase est considérablement activée dans son action par la lumière qui agit ainsi on l'a vu plus haut, de diverses façons pour réduire la teneur des tissus en AIA ; en outre, selon des recherches très récentes, la lumière inciterait les végétaux, par des mécanismes encore mal connus, à élaborer des inhibiteurs d'AIA qui bloquent temporairement son action. On comprend donc bien, grâce à ces rapides indications, que la lumière puisse venir agir, par l'intermédiaire de l'AIA, sur de nombreux processus de la vie des plantes en général et des arbres en particulier.
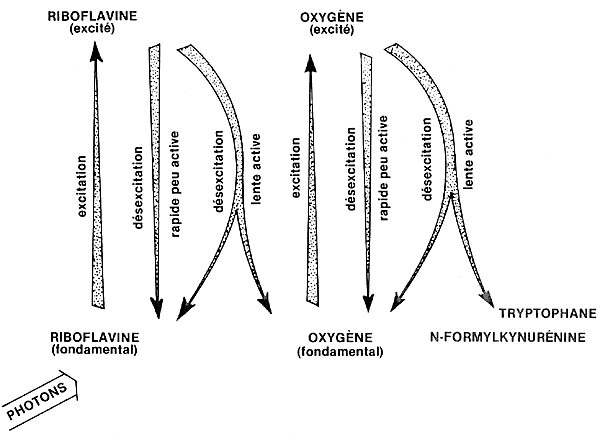
Fig. 2 - Figuration simplifiée du processus de photo-oxydation
du tryptophane. en présence de riboflavine (photosensibilisateur de 1re
classe) et d'oxygène moléculaire; le tryptophane, acide
aminé est considéré comme le précurseur de l'auxine
naturelle (ou AIA), dont il est très voisin chimiquement. La photo-oxydation
provoque l'ouverture du cycle pentagonal, et la transformation en N-formylkyrunenine,
qui ne possède plus les propriétés du tryptophane; ainsi,
la synthèse de l'AIA est arrêtée. (Adapté de
J.-D. Spikes).
La grande activité chimique de l'oxygène excité, sous sa
forme à désexcitation lente, est due en partie au fait que sa
durée de vie peut atteindre plusieurs minutes (et même 1 heure),
alors que les désexcitations rapides ne durent que quelques fractions
de seconde. Par ailleurs, il peut diffuser à des distances relativement
importantes dans les milieux cellulaires.
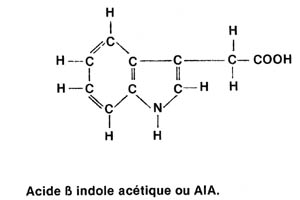 |
Formule développée de
l'AIA Dans le tryptophane, la chaîne 1atérale : CH2-COOH (fonction acide) est remplacée par la chaîne : CH2-CHNH2-COOH (fonction amine et fonction acide), |
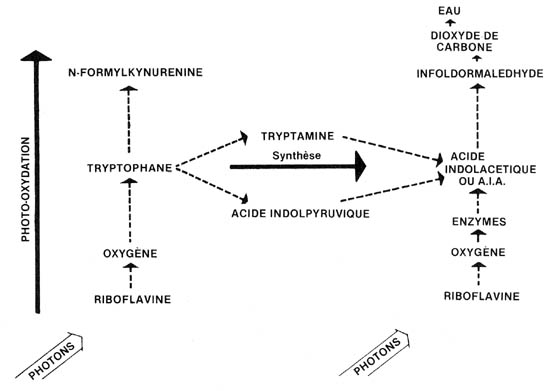
Fig. 3 - Processus probables de la synthèse de l'A.I.A., et de la photo-oxydation de l'A.I.A. et de son précurseur, le tryptophane.
Il existe évidemment d'autres hormones végétales, comme les kinines, les gibbérellines ou des vitamines diverses, mais leur sensibilité à la lumière semble très faible ; quant au pigment photosensible responsable du photopériodisme, le " phytochrome ", son influence paraît surtout évidente pour régler divers processus généraux de la vie des plantes, en liaison avec la durée, variable avec la latitude et avec la saison, du jour et de la nuit. Or, les chênes qui forment des gourmands, et ceux qui n'en forment pas, se développent, côte à côte, sous le même régime photopériodique.
ORIGINE ET DÉVELOPPEMENT DES GOURMANDS ET DES REJETS DE SOUCHES
Les botanistes forestiers s'accordent, en général, pour admettre
avec Ph. Guinier (11) que les gourmands, comme les rejets de souches, ont une
double origine : des " bourgeons proventifs " (ou dormants) qui,
formés en même temps que les autres sur des rameaux au cours de
la première année, ne se développent pas durant les années
suivantes, mais restent en apparence inertes et des " bourgeons adventifs
" qui se forment à un moment quelconque de la vie de l'arbre sur
le tronc ou les racines, et évoluent en pousses aussitôt après
leur formation, d'où leur nom de prompts bourgeons. Sur la tige de l'arbre,
ces bourgeons se forment uniquement sur le bourrelet de recouvrement qui entoure
la section du tronc ou d'une branche. Perrin (6) et Venet (12) ont la même
position.
Alors que la période de latence des bourgeons adventifs est à peu près nulle, comme il vient d'être dit, celle des bourgeons proventifs peut atteindre plusieurs dizaines d'années et l'on assiste à ce phénomène, assez curieux, de petits amas de méristème à activité très réduite, qui se déplacent au fur et à mesure de l'accroissement des arbres en diamètre en restant reliés à la région centrale parfois par une seule file de cellules. Cette lente progression s'arrête au bout de deux ou trois décennies chez certaines espèces comme le bouleau, le tremble, ou le hêtre, mais se poursuit bien plus longtemps chez le charme, le tilleul et le chêne (rouvre et pédonculé) ; puis les bourgeons dormants, qui ne se sont pas développés par absence de " conditions favorables ", disparaissent peu à peu aux regards englobés dans les formations du suber, puis du liber, puis du cambium, enfin du bois.
| Fig. 4 - Région de la Champagne humide; sol de limon des plateaux sur argiles albiennes. Après une coupe de taillis sous futaie, dans l'hiver 1973-1974, état de petites réserves de chêne pédonculé à l'automne 1977 : les gourmands très abondants sur cette espèce sont extrêmement réduits sur les bouleaux et les trembles (à l'arrière-plan). |  |
Quelles sont les " circonstances favorables " qui permettent la reprise
d'activité de ces diverses sortes de bourgeons ? On les voit très
souvent réapparaître et se développer en rameaux, dans les
coupes de taillis-sous-futaie classiques à réserves assez clairsemées,
sur les troncs des futaies de chêne mêmes âgées, et
également sur ceux des petites réserves de charme, de tilleul,
d'érable, d'alisier, etc. maintenues souvent pour conserver aux peuplements
exploités une certaine densité. Sur les chênes, il s'agit
aussi bien de bourgeons proventifs, souvent espacés, reliés à
la partie centrale de l'arbre, que de bourgeons adventifs qui prennent naissance
en bouquets sur les cicatrices des branches tombées précédemment
à la suite du fort ombragement dû aux taillis voisins. Les premiers
donnent, au bout de quelques années, des nœuds plus ou moins importants
dans le bois formé après leur apparition ; les seconds sont à
l'origine des " pattes de chats ", amas de petits nœuds correspondant
aux bouquets de rameaux néoformés. Dans ces deux cas, on dit qu'il
s'agit de " gourmands de lumière ", dont l'origine est souvent
rapportée à l'isolement des cimes des arbres restant sur pied
et à l'afflux de matières nutritives qui en résulte, et
qui " incitent " les bourgeons proventifs à reprendre leur
activité et les bourgeons adventifs à se former rapidement.
Il est à remarquer que les mêmes " circonstances favorables
" se reproduisent au niveau des souches des arbres exploités qui,
très souvent, se garnissent, dès l'année même qui
suit la coupe, de nombreux rejets chez lesquels on peut tout aussi bien discerner
une origine proventive qu'une origine adventive ; là encore, on invoque
l'afflux de sève provenant de la souche et des racines qui, ne pouvant
se répandre dans le surplus de l'arbre disparu, incite les bourgeons
proventifs et adventifs à se développer en des rejets, parfois
puissants, et qui semblent difficiles à contenir. Et cependant, on l'a
dit plus haut, divers forestiers estiment que, si les souches ne sont pas éclairées
d'une façon suffisante, les bourgeons proventifs et adventifs ne se développent
pas.
Un autre type de gourmands, assez curieux, se manifeste souvent dans les coupes
exploitées d'une façon un peu brutale ou bien lorsque, après
une exploitation normale en hiver, les neiges lourdes du début du printemps
viennent recourber un certain nombre de baliveaux : dans ce cas, on observe,
sur la face supérieure de la tige recourbée, le développement
abondant des bourgeons, proventifs surtout, qui forment bientôt une véritable
" brosse " de rejets. Ce fait a frappé également les
spécialistes de la physiologie végétale, qui écrivent
alors " que la courbure de la tige d'un jeune arbre fait disparaître
chez lui la dominance apicale ", ce qui est plus une constatation qu'un
essai d'explication. On retrouve, du reste, le même type de gourmands
sur la face supérieure de troncs rectilignes, faiblement inclinés
(10 à 20° par rapport à la verticale).
Un troisième type de gourmands se manifeste dans des circonstances un
peu spéciales et sur de très nombreuses espèces ligneuses,
en général assez jeune : ce sont les " gourmands d'ombre
" qui se développent sur les troncs des petits arbres surcimés
et d'une végétation générale assez ralentie. H.
Perrin (6) décrit très bien ce genre de gourmands et il estime
même que l'on peut, sans lever la tête, déceler dans un peuplement
les arbres dominés par la seule abondance des gourmands qui se sont développés
sur leurs troncs.
Il est bien évident, dans tous les cas, qu'une fois la levée de dormance des bourgeons proventifs effectuée, et la formation des bourgeons adventifs réalisée, suivie de leur sortie rapide, le développement ultérieur des rameaux retombe sous la dépendance principale de la lumière qui atteint les feuillages, lumière plus ou moins intense selon la position respective de ceux-ci. Il y a donc deux aspects dans ce problème : la naissance des gourmands et des rejets de souches, ET leur développement ultérieur.

Fig. 5 - Région du Barrois ; sol de rendzine calcaire (jurassique) assez superficiel. Coupe effectuée en vue d'un enrésinement, avec une réserve très extensive de charmes. Au bout de 4 à 5 ans, développement très abondant des rameaux latéraux sur tout le pourtour des tiges verticales, et sur la partie supérieure des tiges recourbées.
ESQUISSE D'UNE THÉORIE SUR L'ORIGINE PHOTOBIOLOGIQUE DES GOURMANDS ET
DES REJETS DE SOUCHES
A la base, de ce développement se trouve le " schéma de
Thimann ", dans lequel cet éminent biologiste de l'Université
de Harvard résume le résultat des nombreuses expériences
qu'il fit, avec Skoog, afin de tenter de déterminer les conditions qui
règlent la levée de dormance de certains bourgeons (13) ; évidemment,
cette théorie est déjà ancienne et certains spécialistes
l'estiment dépassée, alors que d'autres continuent à s'y
référer (14). Il est certain qu'elle n'explique pas l'ensemble
des très nombreux et très délicats problèmes relatifs
aux corrélations de croissance, mais, comme on va le voir, sa prise en
considération conduit à un système simple et logique de
représentation des conditions dans lesquelles se forment les gourmands
et les rejets de souches ; en outre, il sera fait une brève mention de
son extension au phototropisme et à " l'effet manchon " (1-2).
L'idée de base retenue par Thimann et Skoog est que le développement de telle ou telle partie du végétal (tige, bourgeons latéraux, racines) est sous la dépendance de la concentration en auxine naturelle (AIA) des tissus susceptibles d'une reprise d'activité, et ces diverses influences sont expliquées par le rapport entre le nombre réel de molécules d'AIA, et le nombre réel " d'accepteurs " existant au niveau de la cellule végétale et variant avec la nature même de l'organe (tige, bourgeon, racine). On a pensé récemment à faire intervenir la proportion entre l'AIA et une autre hormone, la kinine : la différence est minime. Et, comme on l'a dit plus haut, la lumière réduisant de diverses façons la teneur des tissus en AIA, il n'est pas invraisemblable qu'elle puisse, en pénétrant dans la partie externe du tronc de l'arbre, abaisser localement la teneur des tissus en AIA, de telle sorte que celle-ci passe de la concentration " blocage des bourgeons " à la concentration " levée de dormance des bourgeons ".
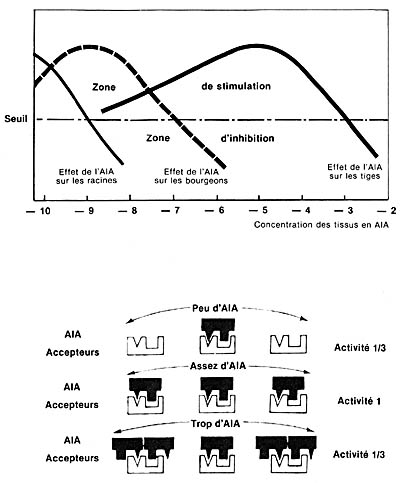 |
Fig. 6 - Partie supérieure : représentation,
d'après K.-V. Thimann, de l'influence de la concentration des tissus
en AIA sur leur développement, selon la nature des parties des
plantes. Partie inférieure : figuration simplifiée des hypothèses qui permettent de comprendre les effets variés de l'AIA suivant son abondance, par rapport aux " accepteurs " existant au niveau cellulaire. Un mode de figuration analogue est utilisé en médecine, pour expliquer comment le même médicament peut avoir, selon sa concentration, des effets très différents sur les organismes humains. |
Voici quelques chiffres qui permettent de préciser mieux cette idée
:
- dans un taillis-sous-futaie dense du nord-est de la France, l'éclairement
horizontal qui atteint les troncs en été dans le milieu
de la journée et par divers types de temps, est " de l'ordre "
de 20 à 50 lux ; après une coupe assez légère (type
: coupe d'abri) laissant 200 à 300 brins et quelques petites réserves
par hectare, cet éclairement passe à 500 / 2 000 lux environ :
il est donc multiplié par 20 à 40 ou 50 (davantage le soir et
le matin, suivant exposition, quand le soleil est bas sur l'horizon) ;
- dans une coupe de taillis-sous-futaie classique, c'est-à-dire assez
forte, l'éclairement latéral passe à 2 000 / 5 000 lux,
donc est multiplié par 100 environ. Quant à la partie supérieure
d'une tige recourbée, il n'est pas rare de mesurer, mais en éclairement
vertical cette fois, des valeurs de 10 000 à 20 000 lux, si le peuplement
restant sur pied est clairsemé.
Or, d'après la théorie quantique qui a été exposée
plus haut, un éclairement de 50 lux correspondrait à un flux approximatif
de 1012 photons par millimètre carré et par seconde
dans la bande des radiations visibles ; si, à la suite d'une éclaircie,
cet éclairement est multiplié par 100, on arrive ainsi à
1014 photons atteignant, dans les conditions indiquées, la
surface externe des écorces. Mais, ces écorces sont-elles absolument
opaques aux rayons lumineux, constituent-elles des obstacles complètement
infranchissables au flux de photons ? Certainement pas, car, grâce à
des appareils de photométrie perfectionnés (les photomultiplicateurs),
il est possible de déterminer que l'écorce d'un chêne, âgé
de 50 à 60 ans, laisse filtrer le 1/1 000 000 environ de la lumière
extérieure. Il passe donc, en moyenne, dans un taillis-sous-futaie dense,
et jusqu'au niveau du liber et du cambium, " de l'ordre " de 106
photons par millimètre carré et par seconde et, à la suite
du suréclairement qui suit une coupe assez importante, ce flux peut s'élever
à 109 ou 1010 photons. Or, d'après les théories
photobiologiques, il suffit d'un ou de quelques photons pour provoquer la photo-oxydation
d'une molécule d'AIA en présence de la riboflavine, et, aux concentrations
observées par Thimann et Skoog, on compte, dans les régions actives
de l'arbre, " de l'ordre " du milliard de molécules de cette
hormone par millimètre cube ; on a dit, par ailleurs, que l'AIA était
élaboré en permanence par les végétaux, et il est
très plausible que, pour chaque intensité d'éclairement,
s'établisse un niveau de concentration " photostationnaire "
de cette hormone, qui favorise le développement plus ou moins important
des bourgeons, jusqu'ici inactifs, et l'apparition des gourmands (*).
Le même type de raisonnement peut, du reste, être fait en ce qui
concerne le traitement en taillis-sous-futaie et les rejets de souches ; il
est certain, en effet, que l'enlèvement de la majeure partie des arbres
d'un peuplement fermé apporte aux souches un afflux de lumière
important qui peut contribuer efficacement, par le même processus, au
développement des bourgeons situés à la base de l'arbre
ou qui naissent sur les bourrelets de recouvrement des souches. On verra, cependant,
que certaines considérations physiologiques laisseraient penser que la
relation est plus complexe.
La question des " gourmands d'ombre " est plus délicate à
interpréter de cette façon ; on peut cependant remarquer que les
arbres dominés, partiellement privés de lumière, ont une
photosynthèse réduite et une activité physiologique générale
diminuée. Il peut en résulter une baisse de l'activité
de synthèse de l'AIA dont la concentration pourrait, par exemple, s'établir
au niveau de 10-8, favorisant, selon le schéma de Thimann,
le développement des branches latérales, au détriment de
celui de la tige principale(**).
(*) Certains lecteurs trouveront peut-être surprenant d'invoquer l'intervention
d'aussi faibles éclairements en physiologie végétale ;
on peut cependant rencontrer des analogies en physiologie animale ; pour expliquer
l'influence de la lumière sur l'activité sexuelle de certains
animaux, on prend en considération les très faibles quantités
de lumière qui pénètrent dans les orifices crâniens
de canards vivants, chez lesquels on a détruit les globes oculaires (J.
Benoît au Collège de France), ou bien qui filtrent au travers des
capsules crâniennes des insectes (Bounhiol à l'Université
de Bordeaux).
(**) Une récente observation de L. Lanier vient, en quelque sorte, corroborer cette interprétation : à la suite de l'été très sec de 1976, sur sol mal alimenté en eau, des perchis de chênes, même (et surtout) très denses, se sont couverts en 1977 de petits gourmands d'origine très probablement proventive. Les " gourmands d'ombre " ne seraient ainsi que des " gourmands de souffrance ".
PREMIÈRES VÉRIFICATIONS EXPÉRIMENTALES (1976-1977)
Pour vérifier dans quelle mesure cette hypothèse correspond à
la réalité, on a d'abord tenté, en 1976, de voir si, en
abritant par des panneaux de plastique noir opaques, des souches de charmes
et de chênes qui avaient été récemment coupés,
on réduirait ou l'on supprimerait même l'apparition des bourgeons
ou leur développement en rejets ; à vrai dire, un argument
théorique semblait vouer cette expérience à un demi-échec,
car il est admis, fréquemment que la teneur en AIA des tissus des végétaux
subit, au niveau du collet, une très forte réduction par suite
de l'intervention de l'auxine-oxydase dont il a été parlé
plus haut. Il le faut bien, du reste, puisque l'on admet, très généralement,
que la teneur en AIA des tissus, propre au développement des racines,
est très nettement inférieure à celle qui favorise le développement
des tiges (très approximativement, 10-10 dans le premier cas,
contre 10-5 dans le second cas). On se trouve donc, a priori, dans
des conditions plutôt favorables au développement des rejets, au
niveau du collet et l'on observe parfois du reste, dans des peuplements de taillis
non exploités mais un peu clairs, l'apparition de telles pousses à
la base des troncs de charme ou de tilleul notamment.
Voici donc, sommairement exposé, ce qui a été organisé et observé, grâce au concours de M. Chevallier, chef de district forestier à Amance. Dans une coupe de conversion située en Champagne humide (affleurement des formations albiennes), exploitée en mars 1976, des panneaux de plastique noir ont été placés, avant le départ de la végétation, sur 7 souches de charme et sur 3 souches de chêne. Au milieu du mois de juillet 1976, alors que les souches voisines non abritées avaient formé, dans ce canton, des rejets nombreux et vigoureux de 0,50 à 0,80 m de hauteur, les souches abritées restaient absolument inertes. Vers la fin du mois de juillet, sont apparues quelques petites pousses, hautes de 0,02 à 0,05 m, complètement décolorées, sur 4 souches de charme et sur une souche de chêne, toutes abritées et elles sont demeurées en cet état jusqu'à la fin de la saison ; la découverte des rejets sur une souche de charme, dès leur apparition, a provoqué leur verdissement rapide, mais sans accroissement appréciable de leur taille. En somme, il résulte de cette petite expérience, qu'en ombrageant très fortement
 |
Fig. 7 - Région de la Champagne humide; sol de limon des plateaux sur argiles albiennes. Etat, à l'automne 1976, de 2 souches de charmes coupés en mars 1976, dans le cadre d'une opération de conversion. À gauche, une souche recouverte aussitôt après l'exploitation, par un panneau de plastique noir (écarté naturellement pour la photographie) est restée complètement inerte, À droite, une souche très proche et de même âge, non recouverte, a formé des rejets nombreux et vigoureux, atteignant 0,60 m de hauteur. |
des souches de chêne et de charme (en fait, un très faible éclairement
pénètre cependant sous les rebords des panneaux de plastique noir),
on supprime ou on retarde considérablement la reprise d'activité
des bourgeons (proventifs et adventifs), lesquels ne donnent, dans le courant
de l'année, que des rejets sans vigueur.
Au cours de la seconde année (1977), les choses restant dans le même
état, on observe d'abord, sur presque toutes les souches recouvertes,
l'installation abondante de colonies de fourmis (ce qui modifie considérablement
les conditions de l'expérience). Les souches de charme sont restées
soit inertes, soit couvertes de quelques très petits rejets décolorés
et en voie de disparition ; par contre, sur les 3 souches de chêne recouvertes,
on a observé quelques rejets assez vigoureux, prenant naissance sur les
" pattes " prolongeant les racines, parfois assez loin du tronc, et,
après une sorte de reptation sous les panneaux de plastique noir, sortant
en dehors de ceux-ci pour pousser ensuite verticalement. On ne peut dire si
cet effet est dû aux très faibles quantités de lumière
qui pénètrent, latéralement, sous les panneaux de plastique
ou à une autre cause.
En ce qui concerne Ies gourmands proprement dits, un dispositif a été installé en 1977 dans une autre coupe de conversion située en Champagne humide et qui venait d'être exploitée ; au milieu du mois de mars, 12 petites réserves de charme, 2 de tilleul, 1 de hêtre et 1 de chêne ont été munies de fourreaux de plastique noir de 1,50 m de haut environ ; comme ce type de coupe est déjà un peu clairiéré, il existait sur ces petites réserves quelques rameaux, en général très minces, datant des années antérieures. Ces rameaux ont été coupés au ras du tronc afin de permettre la mise en place des fourreaux et de vérifier par la même occasion la reprise d'activité à la fois des bourgeons proventifs et des bourgeons adventifs qui pourraient prendre naissance sur les bourrelets de cicatrisation des petits rameaux coupés. Au milieu du mois de septembre 1977, alors que l'on peut considérer l'activité végétative de la partie aérienne des arbres comme à peu près terminée, on a fait les observations suivantes : sur 15 tiges munies de fourreaux noirs, on n'a constaté sous ceux-ci aucune amorce de sortie de bourgeons ou de développement de rameaux latéraux (alors que sur les mêmes tiges, au-dessus des fourreaux,
 |
Fig. 8 - Région de la Champagne humide; sol de limon des plateaux. Coupe de conversion directe des taillis sous futaie classiques en futaies de chêne, exploitée dans I'hiver 1976-1977 ; mise en lumière progressive de semis de chêne pédonculé, en maintenant un léger abri de charme. Les fourreaux de plastique noir sont placés en mars 1977, leur niveau supérieur est à environ 2,20 m du sol. |

Fig. 9 - Région de la Champagne humide; sol
de limon des plateaux. Coupe de conversion directe de taillis sous futaie
en futaie exploitée pendant l'hiver 1978-1977, état à la
fin du mois de septembre 1977. À gauche, perche de charme, non
traitée, présentant de nombreux rameaux latéraux, soit
des années précédentes, soit de l'année même,
jusqu'à la hauteur d'environ 2,20 m du sol (anneau de plastique noir[flèche]).
A droite, perche de charme revêtue, dès le mois de mars 1977 d'un
fourreau de plastique noir jusqu'à environ 2,20 m du sol (anneau de plastique
noir[flèche]), les rameaux latéraux préexistants étant
sectionnés au ras du tronc. Aucune trace de reprise d'activité
des bourgeons, dans toute la partie protégée (le fourreau de plastique
est enlevé, naturellement pour la photographie). Par contre, au-dessus
du fourreau protecteur, activité normale des bourgeons latéraux.
on observait le développement de rameaux, parfois très courts,
et nés cette année même); sur 1 tige de charme (dont la
cime dominée se recourbait sous celle d'un grand chêne), on a observé
sous le fourreau 2 très petits rejets latéraux de l'année,
décolorés, longs de 0,02 et 0,03 m et plaqués le long du
tronc. A titre de comparaison, sur 30 tiges voisines de celles décrites
ci-dessus mais non pourvues de fourreaux noirs, on dénombrait, approximativement,
sur la même hauteur de 1,50 m, 250 rameaux, en général très
minces, nés l'année même ou préexistants des quelques
années antérieures. La privation de lumière, assez poussée
dans ce cas, semblait donc bien avoir retardé considérablement
le développement des bourgeons adventifs et arrêté celui
des bourgeons proventifs ; ceci, pour 1977.
Il aurait été préférable, évidemment, avant
de publier cette étude, d'attendre l'évolution des tiges protégées
par des fourreaux opaques, au cours des années prochaines ; mais on sait
combien sont lents les phénomènes de la végétation
en forêt, et combien il faut prendre de précautions avant de formuler
des conclusions qui n'ont, la plupart du temps du reste, qu'un caractère
provisoire. En réalité, ce genre d'expérience n'est jamais
terminé...
On peut cependant avancer que le " schéma de Thimann ", complété par les indications que donnent les recherches récentes des photobiologistes, permet de " figurer " assez bien les conditions dans lesquelles apparaissent les gourmands (et, moins fidèlement, les rejets de souches) ; à la base de ce phénomène est, évidemment, la notion d'équilibre hormonal, mais ce qui vient d'être dit semble bien établir que la lumière intervient très activement pour modifier, plus ou moins profondément, cet état d'équilibre, et les théories qui viseront, à l'avenir, à donner une interprétation plus fine de ces faits, devront faire intervenir des hormones, ou des inhibiteurs, sensibles à l'action de la lumière ; ceci, si l'on veut qu'elles soient valables pour le milieu naturel.
| Fig. 10 - Représentation de 3 effets différents de la lumière sur les arbres forestiers, en utilisant le schéma général de K.V. Thimann, complété par des considérations photobiologiques. | 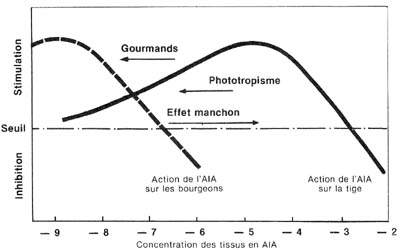 |
Il est à remarquer que le " schéma de Thimann ", complété
comme indiqué plus haut, permet de représenter aussi deux autres
types de photomorphoses, observables de façon courante dans le milieu
forestier : le phototropisme, évidemment, puisque c'est l'étude
de ce phénomène qui est à l'origine même de la découverte
de l'AIA et l'effet manchon (*), c'est-à-dire l'allongement des
tiges de nombreux jeunes arbres quand ils sont soumis à un éclairement
naturel déséquilibré (lumière verticale assez intense
et lumière latérale réduite) (1, 2).
Certains lecteurs penseront peut-être que cet ensemble de considérations est bien compliqué et se demanderont s'il n'est pas possible de faire travailler l'imagination d'une façon plus simple en soutenant, par exemple, que " la lumière excite les bourgeons " selon des processus entièrement inconnus des botanistes non forestiers. C'est évidemment, une façon de procéder, mais n'est-il pas préférable, quand on le peut, de tenter de s'en tenir à des enchaînements d'effets qui s'insèrent dans des " structures d'accueil " solidement établies et qui montrent qu'en définitive, malgré certains caractères originaux, la sylviculture n'est que l'une des applications pratiques de la physiologie végétale classique ?
BIBLIOGRAPHIE
| 1 - ROUSSEL (L.). - Le très jeune épicéa et la lumière. Revue forestière française, n° 1, 1969, pp. 27-32. |
| 2 - ROUSSEL (L.). - De l'emploi des modèles réduits en sylviculture. Revue forestière française, n° 12, 1967, pp. 737-745. |
| 3 - ROUSSEL (L.). - La notion de niveaux d'énergie et son intérêt en sylviculture. Revue forestière française, n° 2, 1970, pp. 131-138. |
| 4 - ROUSSEL (L.). - Production potentielle de quelques types de futaies. Revue forestière française, n° 2, 1971, pp. 273-280. |
| 5 - ROUSSEL (L.). - La formation des gourmands ; essai d'interprétation photophysiologique. - Communication au Groupe d'étude de physiologie de l'arbre, novembre 1973 et Bulletin de la Société forestière de Franche-Comté et des provinces de l'Est, mars 1976, p. 1-6. |
| 6 - PERRIN (H.). - Sylviculture. - Nancy, Ecole nationale des Eaux et Forêts, tome 1, 1952, 318 p. |
| 7 - LEROY-DEVAL (J.). - Les possibilités de traitement de l'okoumé en taillis pour la production du bois papetier. Bois et forêts des Tropiques, n° 161, 1975, pp. 23-34. |
| 8 - GIESE (A.). - Photophysiology. - Londres, Academic press, tome 1, 1964, 377 p. (ouvrage collectif). |
| 9 - TONNELAT (J.). - Biophysique. - Paris, Masson, tome 1, 1968, 402 p. |
| 10 - CENTRE NATIONAL DE LA RECHERCHE SCIENTIFIQUE. - La photobiologie. - Paris, C.N.R.S., 1973, 108 p. (ouvrage collectif). |
| 11 - GUINIER (Ph.), OUDIN (A.), SCHAEFFER (L.), - Technique forestière. - Paris, La Maison rustique, 1947, 376 p. |
| 12 - VENET (J.). - Identification et classement des bois français. - Nancy, Ecole nationale du génie rural, des eaux et des forêts, 1974, 310 p. |
| 13 - THIMANN (K.V.). - The action of hormones in plants and invertebrates. - Londres, Academic press, 1952, 228 p. |
| 14 - CHARTIER (Ph.), BECKER (M.), BONHOMME (R.), BONY (J.-P.). - Effets physiologiques et caractérisation du rayonnement solaire dans le cadre d'une méthode d'aménagement sylvicole en forêt dense africaine, Bois et forêts des Tropiques, n° 152, 1973, pp. 19-35. |