|
Article publié en 1980 dans
la Revue Forestière Française, n° 1, p 35 - (version
.pdf - 584k)
Essences d'ombre
et de lumière
par Louis
ROUSSEL
Il semble que l'on assiste, actuellement, à
une sorte de retour aux conceptions qui, depuis de très nombreuses
années, avaient guidé les forestiers de vieille tradition ;
en effet, de hauts responsables de la politique forestière de notre
pays ont réaffirmé récemment qu'il convenait désormais
d'imiter la nature pour hâter son œuvre et que l'on ne commande
à la nature qu'en lui obéissant. La grande “révolution“
forestière du XXe siècle s'achevant, au sens cinématique
du terme, par une sorte de retour à son point de départ, tout
au moins sur le plan des idées, il importe maintenant d'en développer
les conséquences logiques ; mais, ceci est une autre histoire...
En tout état de cause, pour imiter la
nature, il faut d'abord la connaître et tenter de la comprendre, ce
qui n'est pas toujours une chose facile ; on prendra, comme exemple, une notion
solidement ancrée dans les traditions forestières françaises,
celle de l'existence d'essences d'ombre et d'essences de lumière. Faut-il
considérer qu'elle correspond à une différence réelle
de comportement de ces diverses espèces ligneuses vis-à-vis
du microclimat lumineux, et, dans l'affirmative, est-il possible de formuler
une “explication“ qui soit en accord avec ce que l'on considère
comme acquis, tant en ce qui concerne l'allure de la photosynthèse,
que les processus complexes de la photomorphogénèse ? On s'efforcera
de répondre à ces questions dans les lignes qui suivent.
Il est certain que depuis que les sylviculteurs
interviennent dans les forêts, ils ont remarqué que certaines
espèces ligneuses (comme le Sapin pectiné ou le Hêtre
commun) se sont installées dans des sous-bois,
même très sombres, et qu'elles s'y sont maintenues, tout au moins
pendant quelques années, alors que d'autres semblaient affectionner
plutôt les petites ou les grandes trouées, manifestement mieux
éclairées, tandis qu'elles dépérissaient plus ou
moins rapidement dans les peuplements fermés : le Mélèze
d'Europe, le Pin maritime, les Chênes rouvre et pédonculé,
par exemple, appartiennent à cette seconde catégorie. H. Perrin
(1)*, dans son important traité de Sylviculture, expose ces concepts
avec clarté, en fournissant de nombreux détails, et le lecteur
qui désirera mieux suivre cet article pourra se reporter avec profit
à cet excellent ouvrage. Il y trouvera en particulier un tableau complet,
inspiré de Gayer, dans lequel les espèces forestières
sont classées en essences d'ombre, de demi-ombre, de demi-lumière
et de lumière.
* Les chiffres entre parenthèses renvoient
à la bibliographie.
Mais, parallèlement en quelque sorte à
ce développement des conceptions classiques (« modernisées
» par Wiessner notamment, dès l'année 1908, grâce
à des mesures de l'éclairement régnant dans les diverses
stations forestières), se développaient des idées d'un
genre toute différent, et dont l'un des premiers théoriciens
semble avoir été Fricke, en Allemagne ; cet auteur, dès
l'année 1904, s'élevait assez vivement contre la notion même
d'essences d'ombre et de lumière, « un dogme scientifique
non fondé » selon lui. En privant, grâce à
de petits fossés latéraux, des placettes d'expériences
situées sous divers peuplements, de la concurrence des racines des
grands arbres voisins, il observait une amélioration assez sensible
de la croissance des jeunes Pins sylvestres installés dans ces placettes.
Biquet, dans les Landes de Gascogne, obtenait dès l'année 1912,
des résultats du même genre sur de jeunes Pins maritimes, grâce
à ses « fossés de séquée ». Toumey
aux États-Unis, Hickel en France, Björkman en Suède apportaient
leur appui aux partisans de ce nouveau point de vue, en insistant sur le fait
que l'on devait tenir le plus grand compte de la nécessité d'alimenter
les racines des jeunes sujets en eau et en matières minérales
solubles, ce qui, présenté ainsi, ne soulevait guère
d'objection. C'est, par contre, la tendance à ne considérer
que cet aspect du problème de l'installation et de la survie des espèces
forestières dans les divers sous-bois qui semble excessive, car de
nombreuses expériences, dont certaines sont citées par H. Perrin,
mais dont beaucoup d'autres ont été effectuées plus récemment,
avec contrôle des facteurs énergétiques et hydriques,
ont établi indiscutablement que la lumière intervient d'une
façon très efficace, et différente selon les espèces
en cause, pour déterminer leur installation et leur développement.
Ceci vient confirmer qu'il existe bien, tout au moins pendant les premières
années de leur vie, des espèces forestières d'ombre et
de lumière ; pour adopter une terminologie physiologico-forestière,
on parlera désormais des espèces sciaphiles et héliophiles.
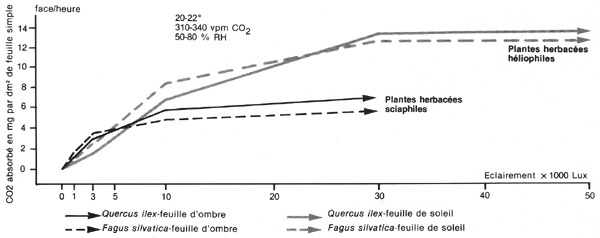
La première idée qui vient à
l'esprit, quand on veut reprendre cette question d'une façon moderne,
est la suivante chez les plantes herbacées, il existe indéniablement
des espèces dont l'activité photosynthétique maximale
est atteinte sous des éclairements faibles ou modérés
(de l'ordre de 5 à 10 ou 20 % par rapport au plein découvert)
et qui se stabilise, aux éclairements supérieurs, à un
niveau moyen ce sont des sciaphiles caractérisées, du type Oxalis
acetosella, par exemple. Par contre, d'autres ont besoin de beaucoup plus
de lumière pour atteindre leur activité photosynthétique
maximale (au moins 40 à 50 % du plein découvert, parfois davantage)
et, à ce moment, la quantité de dioxyde de carbone qu'elles
assimilent, par unité de surface et de temps, est deux fois plus élevée
que celle des sciaphiles ; ce sont des héliophiles, du type nasturtium,
et C. Jacquiot l'expose très clairement dans son ouvrage : La Forêt
(2). On pourrait donc penser que l'on va retrouver la même différence
de comportement entre les espèces forestières considérées
comme sciaphiles, ou comme héliophiles, ainsi qu'il a été
exposé plus haut ; or, très vite, on fait une constatation décevante
: toutes les espèces qui ont été étudiées,
d'une façon précise, dans les appareils modernes à absorption
d'infrarouges (type URAS et IRGA) et dont une
description sommaire fut donnée, dans cette revue même en
des temps reculés (3), présentent, à la fois, des feuilles
d'ombre (à la base et à l'intérieur des cimes) et des
feuilles de lumière (dans les parties les mieux éclairées
des arbres), qui réagissent respectivement de la même façon
que les plantes herbacées sciaphiles et héliophiles. Mais, il
n'est pas possible, de ce point de vue, de déceler des différences
entre la feuille de Hêtre commun et celle du Chêne pédonculé,
ou bien entre l'aiguille de Pin sylvestre et celle du Sapin pectiné.
Du point de vue anatomique, on trouve également le même nombre
de stomates sur la face inférieure de la feuille de lumière
du Hêtre, que sur celle du Chêne (environ 400 par mm2) et les
dimensions des ostioles sont à peu près identiques (7 à
8 de long pour 1 de large, à leur plus grande ouverture).
Au surplus, les physiologistes de l'institut
botanique d'Innsbrück, qui, depuis près de trente ans, ont acquis
une grande maîtrise dans l'étude de la photosynthèse des
grandes espèces forestières (Pisek, Tranquillini, Larcher, Winkler,
Retter et al.) ainsi que des plantes de toutes dimensions, classent dans leurs
publications, les petits végétaux en Schatten-
und Sonnenkräuter, alors
que toutes les espèces forestières, du hêtre au tilleul
et de l'érable au chêne, sont rassemblées en une seule
catégorie, dans laquelle on distingue, évidemment,
l'activité des feuilles d'ombre et celles des feuilles de lumière
(Schatten- und Lichtblätter) (4).
Il faut cependant faire une réserve en
ce qui concerne l'activité photosynthétique maximale du Mélèze
d'Europe (de 50 à 100 % plus élevée, par unité
de surface et de temps) que celle du Pin sylvestre par exemple) ; il
semble, selon des publications récentes, que certaines espèces
du genre Larix ont un « cycle
photosynthétique» intermédiaire entre celui, tout à
fait général, dit en C3 (cycle classique, dit de
Calvin, du nom de son auteur qui reçut en 1961 un Prix Nobel), et celui,
mis en évidence bien plus récemment et à très
fort rendement, dit en C4, réservé à un groupe
restreint de plantes privilégiées (dont plusieurs graminées
tropicales). Les peupliers euraméricains se distinguent également
par leur activité photosynthétique élevée, par
unité de surface foliaire et de temps, mais en maintenant, semble-t-il,
le cycle classique en C3 ; ceci sans doute par suite d'un meilleur
agencement de leur appareil photosynthétique.
On peut rechercher également si, en ce
qui concerne l'éclairement minimal de compensation, on arrive à
trouver certaines différences entre les espèces forestières
sciaphiles et héliophiles. On rappellera brièvement que tous
les végétaux à chlorophylle élaborent, à
la lumière, des substances organiques diverses (glucides, protides
et lipides) qu'ils utilisent pour leur croissance, mais que, pour satisfaire
certains besoins énergétiques, ils en oxydent une partie c'est
le phénomène de la respiration. Les appareils dont il a été
question plus haut permettent d'obtenir des chiffres sur la valeur de l'éclairement
instantané le plus bas, au-dessous duquel la photosynthèse n'équilibre
plus la respiration, et, si cette situation se prolonge, le végétal
meurt, une fois ses réserves épuisées.
On a ainsi mis en évidence que le Hêtre
commun présente encore un bilan positif un peu au-dessus de 300 lux,
alors que le Chêne rouvre doit recevoir au moins 500 lux; le Sapin pectiné
devrait recevoir au moins 300 lux, alors que le Pin sylvestre en exigerait
au moins 1 000. Toutefois, ces indications, obtenues dans des chambres à
atmosphère contrôlée et sur des rameaux convenablement
alimentés en eau, pendant le temps de l'expérience, sont encore
peu nombreuses, et l'on manque souvent de précisions sur la provenance,
l'âge et les conditions de développements antérieur des
sujets des espèces étudiées ; il serait souhaitable,
sans doute, de poursuivre des observations systématiques dans cette
direction.
On insistera davantage sur une autre façon
de considérer le problème des espèces forestières
sciaphiles et héliophiles, et qui a trait, non à la photosynthèse
qui conditionne la nutrition, mais à la photomorphogénèse,
c'est-à-dire à l'influence de la lumière sur la forme
même des très jeunes sujets; comme l'auteur de cette analyse
a déjà publié l'essentiel de ses observations, dans cette
revue même, il se bornera à exposer brièvement des résultats
obtenus, renvoyant, pour plus de détails, aux articles mentionnés
: (5) - (6) - (7) - (8).
Chez les résineux communs en France (espèces
et races étudiées : environ une vingtaine), dès la germination
épigée, on observe une action ralentissante très nette
de la lumière unilatérale, qui se traduit par une courbure phototropique
; ce phototropisme juvénile, qui du reste persiste deux ou trois ans
au moins, est différent suivant les espèces. Très marqué
chez les Pins laricio, sylvestre et maritime, les axes s'orientent parallèlement
à la direction principale des rayons lumineux, dans des cases à
seul éclairement latéral ; pour le Sapin pectiné, races
du Jura et des Vosges, l'axe se maintient vertical sur la majeure partie de
sa longueur, la partie supérieure se recourbant afin de placer les
aiguilles dans un plan à peu près perpendiculaire à cette
direction. L'Épicéa du Jura et des Alpes présente un
type de réaction intermédiaire.
En corrélation, semble-t-il avec ce phototropisme
(les deux types de réactions apparaissent et disparaissent en même
temps), un abri circulaire latéral provoque, chez ces résineux,
un allongement de leur axe hypocotylé et un raccourcissement de leur
radicelle. Mais, comme dans le cas du phototropisme, l'effet est bien plus
marqué chez les espèces considérées comme héliophiles,
que chez les sciaphiles. Si l'on passe aux espèces feuillues, on rappellera
les résultats obtenus depuis une vingtaine d'années sur de jeunes
Chênes, rouvre, pédonculé et rouge d'Amérique,
dont les tiges étaient abritées latéralement par des
manchons opaques plus ou moins larges et qui manifestaient alors un allongement
marqué, par rapport aux témoins non abrités. Dans tous
ces cas, on peut penser qu'au moment de la germination, le très jeune
sujet ne dispose que des réserves de la graine pour « se construire »,
et que ce qu'il utilise pour former son axe aérien et ses premières
feuilles, vient diminuer ce qu'il utilisera pour former sa radicelle ;
cet effet étant particulièrement marqué pour les espèces
à graines légères. Cet ensemble de réactions a
été appelé : « l'effet manchon »
et, dans une publication très récente, l'auteur de cette analyse
en a proposé une interprétation photophysiologique ; la lumière,
surtout latérale, agissant par l'intermédiaire d'un photosensibilisateur
(comme la riboflavine), pour favoriser la photo-oxydation de certaines substances
de croissance, dont l'auxine naturelle, ou AlA.
|
|
|
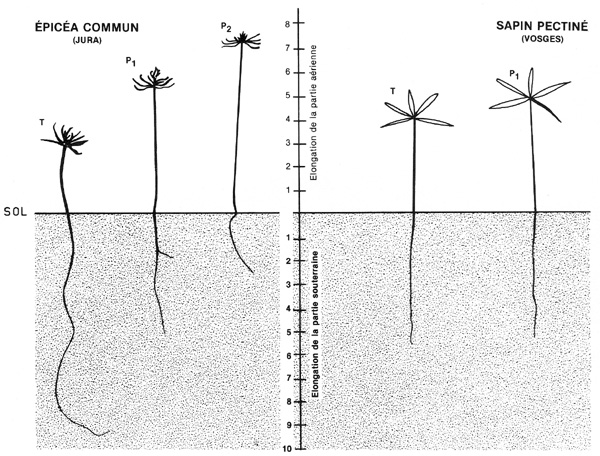
Figure 3
Développement comparé de sujets d'Épicéa
commun et de Sapin pectiné, 3 mois après leur germination,
dans des sols identiques, maintenus à un taux d'humidité
régulier (50 % de leur capacité de saturation).
T :
témoins développés en lumière naturelle.
- P1 : sujets développés en lumière naturelle,
mais abrités de la lumière latérale par un manchon
opaque de 3,5 cm de haut. - P2 sujets développés en lumière
naturelle, mais abrités de la lumière latérale
par un manchon opaque de 7 cm de haut.
|
Il faut remarquer, que dans le milieu forestier,
les choses se passent d'une façon analogue, ainsi qu'on peut l'observer
assez facilement : dans un peuplement un peu dense, la lumière est
principalement dispensée dans une direction verticale, par les interstices
existant dans les cimes, la lumière latérale étant en
général réduite ; dans ce cas, les espèces
sciaphiles allongent peu leurs axes, et raccourcissent peu leurs racines,
alors que les héliophiles s'élèvent le plus qu'elles
peuvent au-dessus du sol, quitte à réduire tellement leurs radicelles
qu'elles n'assurent plus une alimentation suffisante en eau, surtout en présence
des racines des grands arbres en place ; un dessèchement des quelques
centimètres supérieurs du sol, la présence de certaines
substances toxiques (9) peuvent arriver à les faire disparaître,
ou, tout au moins, à réduire leur croissance. C'est de cette
façon, semble-t-il, que l'on peut interpréter les résultats,
exposés précédemment, des expériences de Fricke,
Biquet, et Toumey notamment. À l'inverse, dans les trouées,
les héliophiles se présentent avec un axe hypocotylé
court, donc avec une partie aérienne transpirant peu, et avec une radicelle
longue qui assure mieux leur survie en conditions climatiques défavorables.
Après cette phase de croissance, hypocotylée,
chez les espèces à germination épigée, suit un
bref temps de repos ; puis, très vite, les aiguilles et les feuilles
cotylédonaires se mettent à fonctionner, et la photosynthèse
prend le relais des réserves de la graine, plus ou moins épuisées ;
l'allure de la croissance se modifie alors, surtout pour les espèces
héliophiles. Placées dans des trouées assez bien éclairées,
ayant déjà une radicelle longue (et un axe court), elles se
mettent à s'allonger par les deux extrémités, et la partie
aérienne comme la partie souterraine vont profiter des substances élaborées
par la photosynthèse. Mais, celles qui sont restées dans une
lumière réduite, manifestent une photosynthèse peu active,
et leur forme se modifie peu. Les espèces sciaphiles (résultats
obtenus avec le Hêtre commun et le Sapin pectiné), se maintiennent
mieux, comme on l'a vu, dans les sous-bois même denses (moins de 5 %
d'éclairement relatif, par exemple), mais, faute de lumière,
leur allure générale ne change guère ; celles qui se
sont installées dans les trouées, mieux éclairées,
et manifestant une activité photosynthétique plus marquée,
vont utiliser les substances qu'elles élaborent à allonger et
à ramifier leurs racines, pendant que leur partie aérienne s'épaissira,
mais ne s'allongera guère.
Tous ces problèmes ont été
également étudiés en Italie, spécialement à
l'institut d'écologie forestière et de sylviculture de l'Université
de Florence, notamment par Giannini, dès l'année 1971 (10) -
(11) ; voici quelques chiffres démonstratifs, se rapportant du reste
à un essai de 98 jours, à partir de la germination, c'est-à-dire
que les résultats obtenus englobent les deux phases de croissance,
hypo- et épicotylée, définies plus haut. Dans l'un de
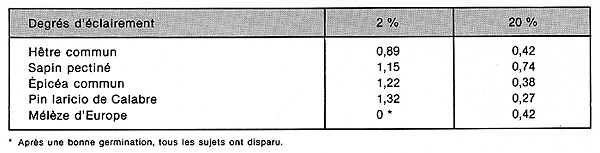
ces essais, 6 espèces ont été observées sous 3
degrés d'éclairement relatif (obtenus grâce à des
filets de plastique « umbratex »), soit 2 % 20 % et
45 %. On donnera seulement les chiffres relatifs aux degrés 2 % (peuplement
très dense) et 20 % (petite trouée), qui sont les plus démonstratifs.
Rapports des longueurs des
parties aériennes et souterraines
|
|
Figure 4
Pourcentage de survie, au bout de 98
jours, des semis de diverses espèces ligneuses,
1 feuillue et 4 résineuses, sous
3 éclairements relatifs, d'après les expériences
de Giannini (1971).
|
La longueur moyenne de la racine (au bout de
98 jours) s'accroît, chez le Hêtre commun, quand on passe de l'ombre
dense à un éclairement moyen, de 48 % environ, chez le Sapin
pectiné de 75 %, mais chez le Pin laricio de Calabre de 337 %. On doit
remarquer cependant que ces expériences sont faites sur des races d'espèces
se développant dans l'Italie centrale, donc considérées,
en général, comme plus héliophiles que celles du Nord-Est
de la France ; aussi est-ce avec beaucoup d'intérêt que
l'on a accueilli les résultats, publiés en 1978 (11), de nouvelles
expériences concernant le Sapin pectiné, sous 7 intensités
d'éclairement relatif : 1 % ; 3 % ; 9 % ; 15 % ; 33 % ; 50
% ; et 100 %, et étendues sur 2 années. En quelques mots, on
indiquera que le dispositif comportait une série d'essais sur sol sec,
et une autre sur sol humide ; sous tous les éclairements relatifs,
la germination des graines a été bonne, mais, sous 1 % de lumière
relative les sujets ont rapidement disparu, sous 3 % assez peu existaient
encore à la fin de la seconde année, les meilleurs résultats
étant obtenus entre 15 et 33 % ; la hauteur de la partie aérienne
des sujets était à peu près la même, sous tous
les degrés d'éclairement permettant la survie des sujets, mais
la longueur des racines augmentait avec l'intensité de la lumière.
La conclusion des auteurs de cette expérience est que le Sapin des
Apennins est une race « tolérant l'ombre » ; elle
vient confirmer d'une façon très intéressante la position
de Giacobbe, éminent spécialiste de l'écologie et de
la sylviculture méditerranéennes, qui dans une importante étude,
traduite intégralement par l'institut de botanique de Montpellier,
a récemment disserté sur ce qu'il appelle : « la
pseudo-héliophilie » des espèces ligneuses de l'Italie
centrale (12).
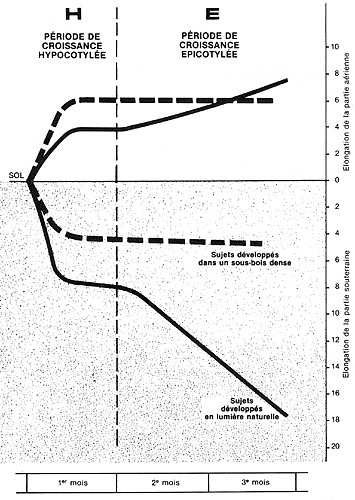 |
Figure 5
Représentation approximative du développement en longueur
de la partie aérienne et de la partie souterraine d'une espèce
héliophile typique (Pin laricio noir d'Autriche) pendant les 3
mois qui suivent sa germination.
N.B. Dans le cas d'une espèce sciaphile (type Sapin pectiné),
les courbes sont bien plus rapprochées les unes des autres. |
Que conclure de cet exposé, certes un
peu touffu, mais la physiologie végétale n'est-elle pù,
elle-même, une discipline complexe ?
Il existe très probablement des espèces
forestières sciaphiles et héliophiles, tout au moins dans les
parties extrêmes de la classification de Gayer-Perrin.
On ne trouve guère de différences
entre l'activité photosynthétique des feuilles ou des aiguilles
de ces deux sortes d'espèces, prises dans des positions homologues
des cimes.
Un indice pourrait sans doute être trouvé
dans la recherche de l'éclairement relatif de compensation.
L'élément qui semble le plus démonstratif
est le changement de forme des très jeunes sujets, qui conditionne
leur survie : chez les sciaphiles, dans les sous-bois denses, chez les héliophiles,
dans les trouées mieux éclairées.
Mais de nombreux aspects de ces problèmes
demeurent encore obscurs, et, fort heureusement pour les chercheurs forestiers,
le sujet n'est nullement épuisé.
BIBLIOGRAPHIE
1 - PERRIN (H.). - Sylviculture. Nancy, École
Nationale des Eaux et Forêts, tome I, 1952, 318 p.
2 - JACQUIOT (C.). - La Forêt - Paris,
Masson, 1970, 160 p.
3 - ROUSSEL (L.). - Le
Phytotron du Patscherkofel. Revue Forestière Française,
n° 12, 1960, pp. 769-774.
4 - LARCHER (W.). - Okologie der
Pflanzen. Stuttgart, Ulmer, 1973, 320 p.
5 - ROUSSEL (L.). - Le
phototropisme juvénile des résineux. Revue Forestière
Française, n° 11, 1966, pp. 708-717.
6 - ROUSSEL (L.). - De
l'emploi des modèles réduits en sylviculture. Revue Forestière
Française, n° 12, 1967, pp. 737-745.
7 - ROUSSEL (L.). - Le
très jeune épicéa et la lumière. Revue Forestière
Française, n° 1, 1969, pp. 27-32.
8 - ROUSSEL (L.) - Lumière,
gourmands et rejets de souches. Revue Forestière Française,
n° 3, 1978, pp. 186-200.
9 - DUCHAUFOUR (Ph.)
et ROUSSEAU (L.Z.). - Les phénomènes d'intoxication des plantules
de résineux par le manganèse dans les humus forestiers. Revue
Forestière Française, n° 12, 1959, pp. 835-847.
10 - GIANNINI (R.). - Ricerche
sull'azione della luce nei confronti della sopravvivenza e dello sviluppo
di semenzali di varie specie forestali. Accademia Italiana di Scienze Forestali,
1971, pp. 201-225.
11 - GIANNINI (R.). - TASCIONE
D. - Ulteriori indagini sull'influenza della luce sulla nascita e le sviluppo
di semenzali di abete bianco. L'Italia Forestale e Montana, n° 5 1978,
pp 213- 225.
12 - GIACOBBE (A.). - Ricerche
sull'eliofilia delle foreste nel clima mediterraneo. Accademia Italiana di
Scienze Forestali, 1975, pp. 251-275